【連載:数理生物学の生命観 #4】現象と数理モデルの双方向性がもたらす美しさ

【連載:数理生物学の生命観】
#1 研究者と芸術家の共通点を探る
#2 花をモデルにしたグラフィック作品を制作する芸術家の視点から
#3 生命を扱うアーティストが提示する「感得者を含むモデル」の可能性
#4 現象と数理モデルの双方向性がもたらす美しさ
#5 生命システムの数理 – 現象の背後にある普遍性を目指して
#6 「モデリング」で芸術と科学を再融合する
2019年9月に数理生物学会が開催したシンポジウム「数理生物学の生命観」の講演内容を紹介する本連載。今回ご登場いただく山道真人氏は、集団遺伝学と数理モデルの知識を活かし、群集・生態系における生態ー進化相互作用について、フィールドワークも交えつつ精力的に研究を行なっています。
本記事では湖沼群集のフィールド実験のデータを数理モデルに落とし込む過程を説明していただきました。実際のデータを数理モデルで表現できることへの美しさについても踏み込んだ内容となっています。
研究で感じる3つの美しさ
私は生態学・進化生物学といった、マクロなスケールの生物学について、おもに数理モデルを用いて研究を行っています。このような研究を行ううえで感じる美しさには、大きく分けて3つあると考えています。
1つ目は、数学的な美しさです。下図左のような、単純で普遍的な数式によって、複雑で多様な自然現象の背後に潜む統一的なルールを記述できるということに対して、美しいと思う感覚があります。
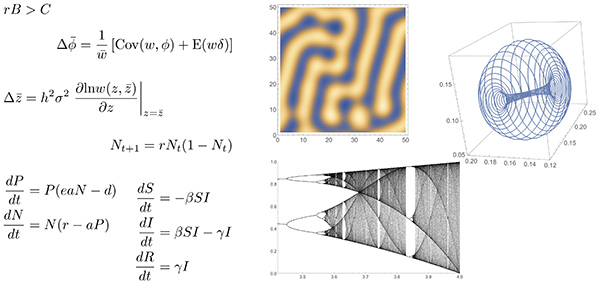
2つ目は、数理モデルのシミュレーションの結果として現れる複雑なパターンに対して感じる、視覚的な美しさです。上図右のような、力学系のアトラクター、カオスの分岐図やチューリング・パターンは、その背後のモデルの単純さも相まって美しく思われます。
3つ目は、数理モデルから実際の生物現象を予測し、予測に基づいて実験を行い、そこで得られたデータに見られる複雑なパターンを、数理モデルによって説明するといった、理論と実証の双方向性です。これについては次のセクションで詳しくご紹介します。
現象と数理モデルの双方向性:遮光された湖沼群集を例に
生態学・進化生物学のようなマクロ生物学では、対象とする時間・空間のスケールが大きく、多様な要素が複雑に相互作用するため、観測された生物現象を理解することが難しい場合が多いです。そのため、ロジックを積み重ねて、複雑なフィードバックを考慮することのできる数理モデルが非常に有用になります。理論研究と実証研究が連携することで、これまで数多くの重要な結果が得られています。
このような理論と実証の双方向性に関する例として、最近行った研究を紹介します。この研究は、東北大学の占部城太郎教授のプロジェクト(科研費基盤研究A「水界生物群集に及ぼす光―栄養バランスの生態化学量効果:北米での野外実験による検証」)の一環として、米国ニューヨーク州のコーネル大学が所有する実験池で行ったものです。
マンハッタンから車で4時間ほど北西に行ったイサカという小さな町にあるコーネル大学は、30メートル四方、深さ1.5メートルの研究用実験池を60個ほど所有しています。コーネル大学の陸水学者のNelson G. Hairston Jr.教授(現名誉教授)と共同で、このうちの6個の池を借りて、家庭用プールでカバーとして使う円形のシートを大量に購入し、水面に浮かべました。このように生態系の要である太陽光が遮られたときに、湖沼の生態系はどのように反応するのでしょうか?
占部教授が過去に行った研究では、光が強いときには植物プランクトンが光合成を活発に行うことで、炭素過多になり、餌としての質が低くなる(ミジンコなどの動物プランクトンにとってリンを摂取しにくくなる)一方で、光が弱い時には光合成が十分に行えないため、中程度の光で動物プランクトンの増殖率が最大になるという実験結果が得られ、数理モデル解析でも確かめられてきました。
しかし、これまでは池をまるごと使ったスケールの実験は行われていませんでした。この野外実験では、池の水面を8割近く覆った暗い池、5割ほど覆った中程度に暗い池、遮光しないコントロール(対照実験)の池という3つの処理を作り、生物群集の変化を比べました。

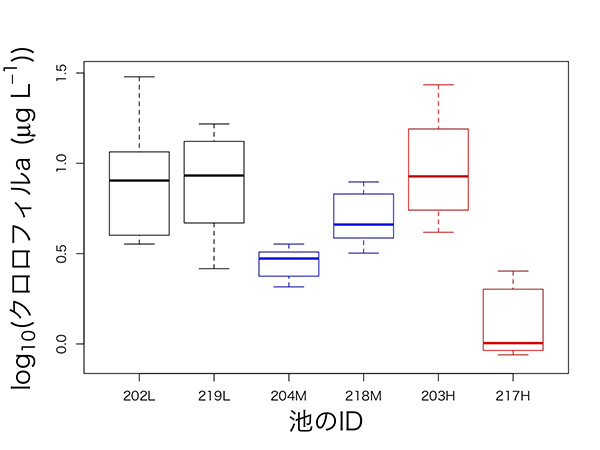
風間健宏博士(現国立環境研究所)が中心となって、2015年の6月から9月まで、2週間ごとに6つの池で植物プランクトンの個体数の指標となるクロロフィルaの量を測ったところ、下図のような時系列データが得られました。複雑でわかりにくいパターンに見えますが、初期値を除いて箱ひげ図で表すと、何となくパターンが見えてきます。
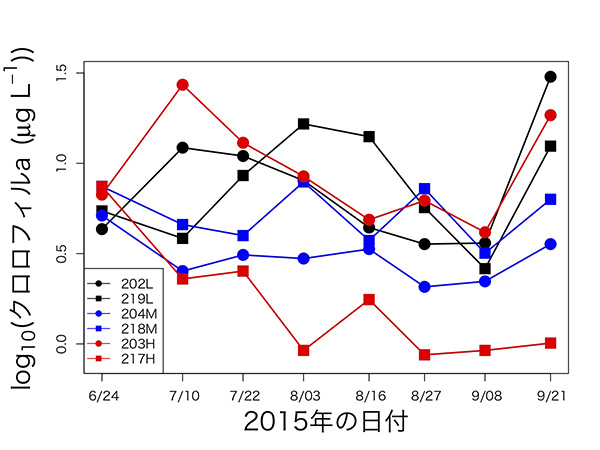
203Hという池を除いて考えてみると、赤で示した明るい池(217H)でクロロフィル量が最も少なく、黒で表した暗い池(202L・219L)では最も多く、青い中程度の明るさの池(204M・218M)では、中間的な結果になっています。つまり、「池を暗くすると光合成ができなくなって、植物プランクトンが減るはず」という当初の予想に反して、池が暗いほど植物プランクトンが増えたという意外な結果が出てきました。
このようなパターンが明らかになってきた8月に、各実験池の底に生えている水草(シャジクモ等)の量を測りました。池の底の泥を取ってきて、その泥の表面にどれくらい水草が生えているか定量すると、クロロフィル量と負の相関があることがわかりました。この結果から、植物プランクトンと水草は何らかの形で競争しているために負の相関が出るのではないかと考えました。それでは、全体のパターンから外れている、遮光していない203Hの池では、何が起きているのでしょうか?
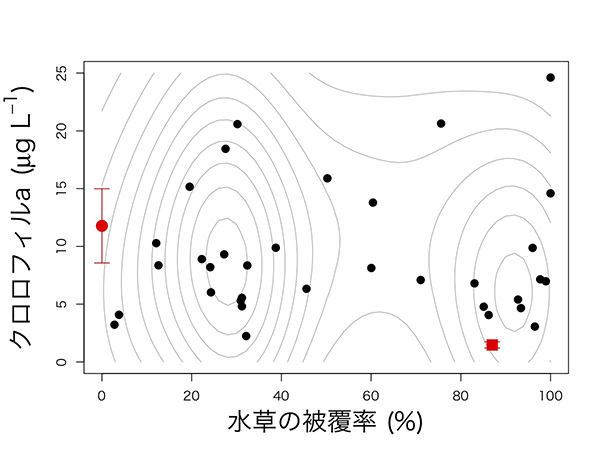
遮光していない池で何が起きているのかを理解するために、実験に使っていない35個の池を調べました。遮光実験を行った2015年の翌年(2016年)にドローンを飛ばして、池の上空から写真を撮り、池の底がどれくらい水草に覆われているか、定量化しました。

結果を下図に示します。縦軸がクロロフィルの量、横軸が水草の被覆率で、黒い点が2016年の35個の池の結果、赤い点が2015年の2つのコントロールの池です。これを見ると2015年のコントロールの池は、ふた山型になっている全体の分布を反映した結果のように見えます。この背後にあるメカニズムを考えるために、数理モデルの解析を行いました。
数理モデルとしては、池の中の植物プランクトンと水草が、光と栄養塩をめぐって競争している状況を考えました。水面の上から入ってくる太陽光は、まず植物プランクトンによって吸収され、その残りを池の底に生えている水草が吸収するため、光をめぐる競争では植物プランクトンが有利になります。その一方で、栄養塩は池の底に含まれるために、まず水草が吸収して、その残りを植物プランクトンが吸収すると仮定しました。式にすると下図のようになり、非常に複雑です。パラメータが多いですが、植物プランクトンと水草の増殖について調べた先行研究からパラメータ値を引用しました。
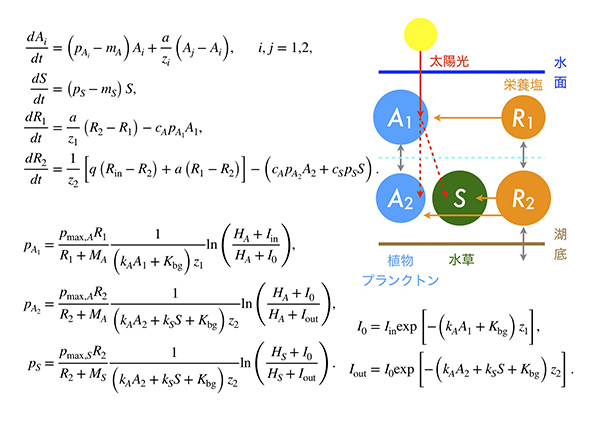
数理モデルのシミュレーションを行うと、下図の左のような結果が得られました。aの縦軸は植物プランクトンの量で、cの縦軸は水草の量です。横軸はどちらも太陽光の量です。光が少ない状況では、池の底の水草まで光が届かなくなって植物プランクトンが勝ち、水草は絶滅します。一方、光が強い状況では、栄養塩をめぐる競争が強くなり、植物プランクトンは絶滅して、水草が勝つということがわかりました。
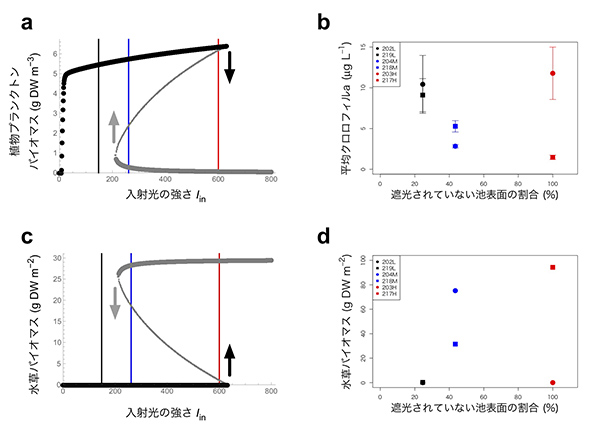
光の強さが中程度の時には、「双安定性」が起きます。ここでは、初期値に応じて、(1) 水草が優勢で植物プランクトンがほとんどいない状態か、(2) 植物プランクトンが優勢で水草が絶滅している状態のどちらかになるということが数理モデルから予測できました。実は浅い湖では、多数派が有利になるために、このような双安定性が起こることが、過去のさまざまな研究から明らかになっていました。しかし、過去の研究では栄養塩の量と双安定性を調べた研究が多く、光の量はあまり注目されてきませんでした。この数理モデルの結果(a, c)を、実際のデータ(b, d)と比べると、
(1) 光が強い(コントロールの)池では、双安定性があり、植物プランクトンが多くて水草が少ない状況か、植物プランクトンが少なくて水草が多い状況になる
(2) 光が弱まった(5割遮光の)池では、(1)の植物プランクトンが少なく水草が多い池よりも、植物プランクトンの量が増え、水草が減る
(3) さらに光が弱い(8割遮光の)池では、植物プランクトンがさらに増え、水草はほとんどいなくなる
というパターンが合致していることが見えます。そのため、複雑に見える野外大規模実験の結果に対して、数理モデルを用いてもっともらしい説明を与えることができた、と言えるかと思います。ただし、遮光と湖沼群集の変化の間の因果関係を完全に証明するためには、より詳細にコントロールされた実験を積み重ねていくことが必要です。
最近では、ソーラーパネルをため池等に浮かべて発電を行う水上太陽光発電が広まりつつあります。私たちの論文では、浅い池にソーラーパネルを浮かべて遮光した場合、水草が減って植物プランクトンが増え、池の水質が悪化するといったことも起こりうるので、注意が必要ではないかという議論をしています。
数式という媒介を通じて生命をいかに表現可能か?
再び今回のシンポジウムの内容に立ち返ると、数理生物学の研究者は数式という媒介を通じて、生命現象をいかに表現可能かという問題に常に直面しています。しかしその一方で、数理モデル研究の長い歴史の積み重ねを経て、ある程度決まった作法からモデル化することに慣れてしまっているという側面もあります。
生態学や進化生物学では、100年程前にダーウィンが著した『種の起源』のような数式を用いない言葉による「理論」から、生態学のロトカ・ヴォルテラ、集団遺伝学のライト・フィッシャー・ホールデンらによる数理モデル解析の発展を経て、現在では実験・観測と並ぶ重要な科学的アプローチのひとつとして、数理モデル解析が精緻化していく流れにあると感じています。今回のシンポジウムをきっかけに、「アート」としての数理モデルを考え直してみるのは、今後の多様な数理モデル研究の発展のために重要な点だと感じています。
参考文献
- 加藤元海 (2005) 生態系における突発的で不連続な系状態の変化:湖沼を例に. 日本生態学会誌 55(1): 199-206 DOI: 10.18960/seitai.55.1_199
- Loladze I, Kuang Y, Elser JJ (2000) Stoichiometry in producer-grazer systems: linking energy flow with element cycling. Bulletin of Mathematical Biology 62: 1137-1162 DOI: 10.1006/bulm.2000.0201
- Urabe J, Kyle M, Makino W, Yoshida T, Andersen T, Elser JJ (2002) Reduced light increases herbivore production due to stoichiometric effects of light/nutrient balance. Ecology 83(3): 619-627 DOI: 10.1890/0012-9658(2002)083[0619:RLIHPD]2.0.CO;2
- Urabe J, Sterner RW (1996) Regulation of herbivore growth by the balance of light and nutrients. Proceedings of the National Academy of Sciences of the United States of America 93(16): 8465-8469 DOI: 10.1073/pnas.93.16.8465
- Yamamichi M, Kazama T, Tokita K, Katano I, Doi H, Yoshida T, Hairston NG Jr, Urabe J (2018) A shady phytoplankton paradox: when phytoplankton increases under low light. Proceedings of the Royal Society B: Biological Sciences 285(1882): 20181067 DOI: 10.1098/rspb.2018.1067
- 吉田丈人 (2007) 「理論と実証」企画趣旨. 日本生態学会誌 57(2): 172-173 DOI: 10.18960/seitai.57.2_172
講演者プロフィール
山道真人
学部ではフィールド科学としての生態学を、大学院では理論的な進化生物学を学び、現在は豪州クイーンズランド大学でマクロ生物学、特に迅速な進化と個体群・群集動態のフィードバックについて、おもに数理モデルを用いて研究している。
この記事を書いた人
-
切江志龍
東京大学農学生命科学研究科博士課程在籍。主に植物フェノタイピングと形態モデリングの研究を行っています。生物・生命にまつわる芸術や文化史にも興味を持ち、修士課程からは画家モネの描いたスイレンの園芸史について調査しています。
堀部和也
大阪大学大学院理学研究科博士課程。主に大脳皮質の形づくりについてコンピュータシミュレーションを用いた研究を行っている。仮想生物の進化をコンピュータ上で再現し、生まれてくる新奇な個体の観察が日課。生物・生命の理論モデルからそれらを対象とする芸術にも関心を持ち、ハッカソンで出会ったアーティストと一緒に制作を行っている。
この投稿者の最近の記事
 研究成果2020年10月19日【連載:数理生物学の生命観 #6】「モデリング」で芸術と科学を再融合する
研究成果2020年10月19日【連載:数理生物学の生命観 #6】「モデリング」で芸術と科学を再融合する 研究成果2020年10月19日【連載:数理生物学の生命観 #5】生命システムの数理 – 現象の背後にある普遍性を目指して
研究成果2020年10月19日【連載:数理生物学の生命観 #5】生命システムの数理 – 現象の背後にある普遍性を目指して 研究成果2020年10月19日【連載:数理生物学の生命観 #4】現象と数理モデルの双方向性がもたらす美しさ
研究成果2020年10月19日【連載:数理生物学の生命観 #4】現象と数理モデルの双方向性がもたらす美しさ 研究成果2020年10月19日【連載:数理生物学の生命観 #3】生命を扱うアーティストが提示する「感得者を含むモデル」の可能性
研究成果2020年10月19日【連載:数理生物学の生命観 #3】生命を扱うアーティストが提示する「感得者を含むモデル」の可能性


