ゼニゴケにはあってタバコにはない遺伝子を追求したら、100年越しの謎が解明できた – 名古屋大・藤田祐一教授インタビュー【前編】
植物にとって、最も重要な栄養成分のひとつである窒素。空気中には大量に含まれるありふれた元素だが、植物の生育にはそのままでは利用できないため、現状では肥料として補給する必要がある。しかし、将来的に空気中の窒素を自ら肥料に変換して生育する植物が実現できれば、作物生産に伴う環境・エネルギー負荷を大幅に軽減できるかもしれない。そんな夢のような「窒素固定作物」実現のカギとなるのが、ニトロゲナーゼという酵素だ。
ニトロゲナーゼは、一部の原核生物だけが備えている酵素で、大気中の窒素をアンモニアに変換する能力をもつ。名古屋大学大学院生命農学研究科 藤田祐一教授は、ニトロゲナーゼを植物でも機能させるというアプローチで、窒素固定作物の創出を目指している(参考:academist Journal「空気を肥料とする植物は可能か? – 光合成生物で窒素固定酵素を作動させる試み」 )。
実は、藤田教授とニトロゲナーゼの付き合いは長く、出会いは大学院生時代にまで遡る。本インタビューでは、藤田教授の大学院生時代の研究と窒素固定作物創出に向けた研究とのつながり、そして窒素固定作物実現までの展望を、前後編の2本に分けてお届けしたい。

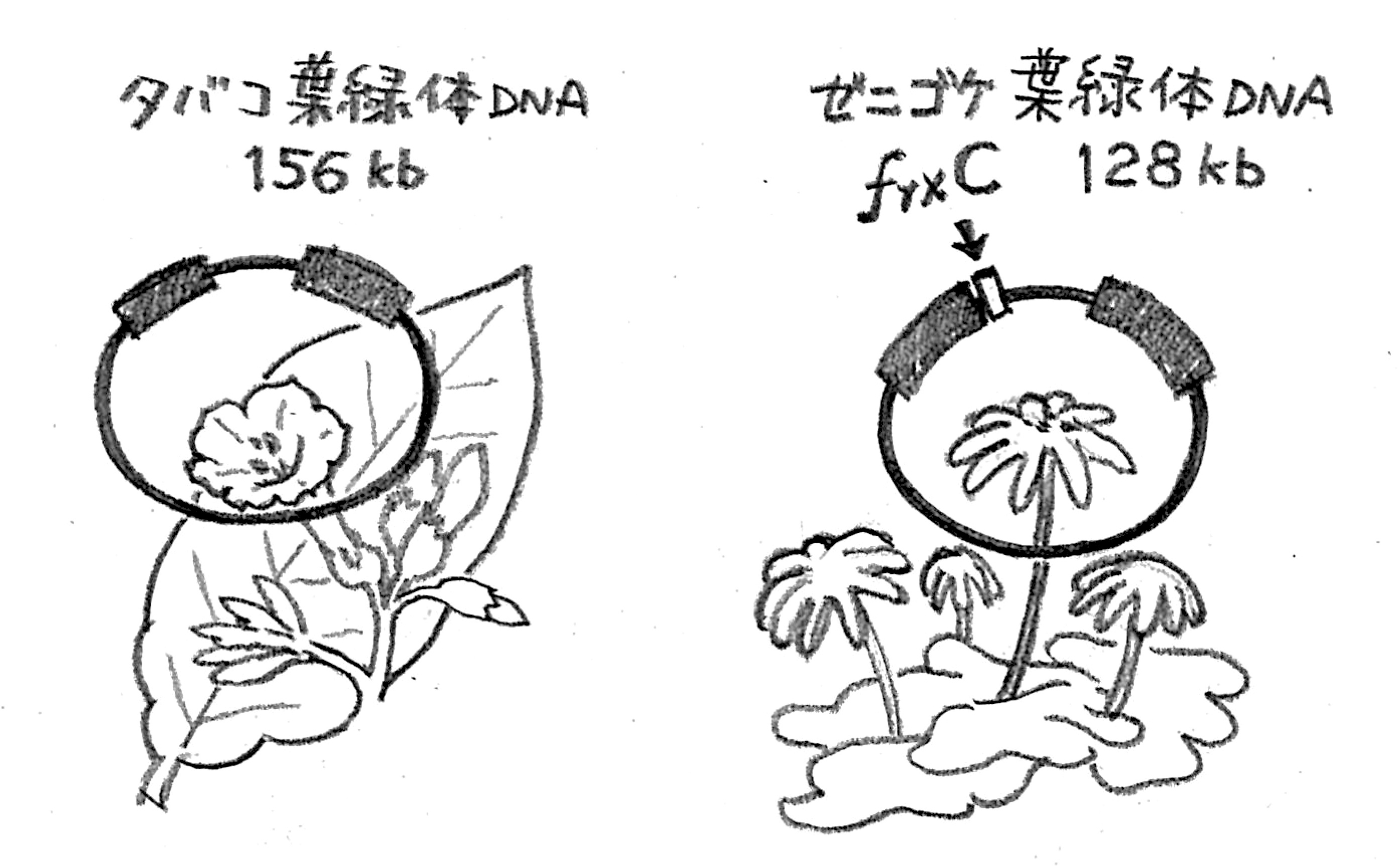
ゼニゴケ葉緑体にはあって、タバコ葉緑体にはない謎の遺伝子frxC
——以前academist Journalにご寄稿いただいた研究コラムで、大学院生の頃にニトロゲナーゼと類似したタンパク質の機能を明らかにしようとする研究を始めたと書かれていましたね。まずは、藤田先生の大学院生時代の研究について教えていただけますか。
大学院生時代には、ゼニゴケの葉緑体DNA中にある機能未知の遺伝子について研究していました。日本の研究グループが世界に先駆けてゼニゴケとタバコの葉緑体DNAの全塩基配列解読に成功したのは、1986年のことでした。当時私はちょうど修士課程1年で、指導教員から、ゼニゴケ葉緑体DNAの中に何をしているのかわからない不思議な遺伝子があるのだが、その機能を調べることを新しい研究テーマにしてはどうかと提案されました。frxCと名付けられていたその遺伝子のアミノ酸配列を調べると、ニトロゲナーゼを構成するひとつのサブユニットによく似ているんです。ゼニゴケは立派な真核生物なのに、原核生物しか持っていないはずのニトロゲナーゼによく似た遺伝子がなぜ葉緑体DNAに存在しているのか、確かに不思議ですよね。そこで、frxCの機能解明に向けた研究をスタートすることにしたんです。
ただ、機能解明に向けた実験を行ううえで何を調べたらその手がかりになるのかわからなくて困りました。なにしろアミノ酸配列がニトロゲナーゼに似ているということしか情報がなかったので、まずはニトロゲナーゼについて知るしかないですよね。というわけで、ニトロゲナーゼのことは大学院生時代にかなり勉強していました。また実は、現在の窒素固定作物の研究で使っている窒素固定性シアノバクテリアも当時の研究で利用し始めたものです。frxCの機能を調べるにはfrxC遺伝子を欠損した変異株を解析する必要がありますが、当時ゼニゴケでfrxCの欠損株をつくることは困難でしたので、葉緑体のモデルとしてこのシアノバクテリアを使うことにしたんです。
——ニトロゲナーゼと窒素固定性シアノバクテリアとはその当時からの縁なんですね。
1990年(博士課程3年生の秋頃)には窒素固定性シアノバクテリアの形質転換系の確立に成功し、frxC欠損株を単離し、その形質解析を行いました。形質を調べていくなかで、暗所で培養した野生株は緑色をしているのに、frxC欠損株は青いことに気が付きました。暗所で培養した野生株は緑色の色素であるクロロフィルを含むので緑色に見えますが、frxC欠損株ではクロロフィルが合成されなくなったことで青くなってしまっていたんです。実際に吸収スペクトルを見ても、クロロフィルが合成されていないことは明らかでした。
みなさんよくご存知の「ガリガリ君 ソーダ味」の青色は、シアノバクテリアがもっているフィコシアニンという色素(スピルリナ青という食用色素)がもとになっています。クロロフィルがなくなると、このフィコシアニンの青色が残って青く見えるんです。

さらに解析を進めると、frxC欠損株では、クロロフィルの代わりにプロトクロロフィリドという物質が蓄積されていることもわかってきました。クロロフィル生合成の最終段階では通常、プロトクロロフィリドがクロロフィルの直接の前駆体であるクロロフィリドに変わる反応が起こります。プロトクロロフィリドは淡い黄緑色の色素ですが、分子内の1箇所の二重結合が単結合に変わる還元反応によって、ポルフィリン環の電子の共鳴状態が変わることで、クロロフィルと同じ鮮やかな緑色を示すようになるんですよね。
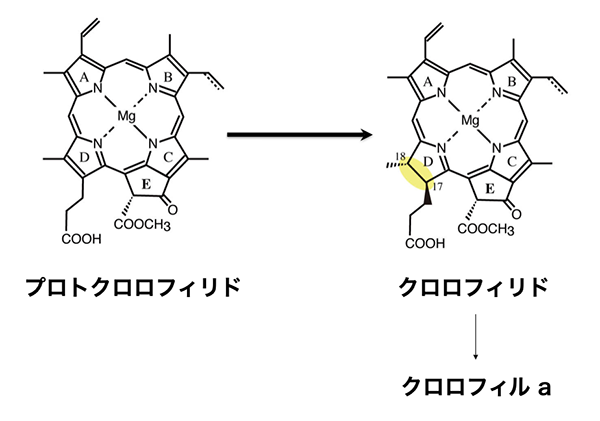
プロトクロロフィリドが蓄積してしまっているということは、frxC欠損株ではこのクロロフィル生合成反応の最終段階が起きていないということを示しています。さらに不思議なのが、クロロフィル生合成反応が止まってしまうのは、暗所のみであるということ。明るいところでは、frxC欠損株でも普通にクロロフィルをつくるんです。
——frxC欠損株は、明るいところではクロロフィルをつくるのに、暗いところではクロロフィルをつくる反応を行わないということですね。なぜそのようなことが起きるのでしょうか。
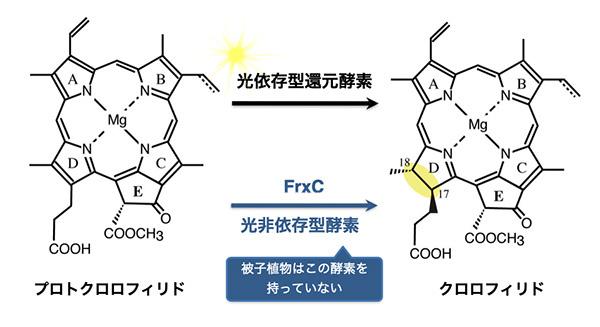
多くの植物は、プロトクロロフィリドをクロロフィリドに変換する酵素として光に依存するタイプの酵素(光依存型還元酵素)しかもっていません。みなさん、暗いところで植物を育てると、必ずもやしのように黄色っぽくなってしまうと思っていませんか? 確かに被子植物は暗いところで育てると緑色にはならずもやしのようになります。でもそれって実は、光合成生物全体から見るとあまり普通なことではないんです。
クロマツなど裸子植物を暗所で芽生えさせると、クロロフィルをつくってちゃんと緑色になります。今から100年以上前の1900年にドイツの植物生理学者が、さまざまな裸子植物を明所と暗所で芽生えさせる実験を行い、クロマツの芽生えが緑色になるという観察結果を報告しています。しかし、被子植物と裸子植物とでなぜこのような違いが出てくるかということは、誰もわかっていませんでした。実は、frxCの機能がこの謎に関わっていたんです。
被子植物では、光依存型還元酵素のはたらきにより、クロロフィルの生合成が行われます。つまり、光がないとこの反応は進まないので、暗所で育てると、被子植物はクロロフィル生合成がプロトクロロフィリドの段階で止まってしまってクロロフィルをつくることができません。
しかし、シアノバクテリアや裸子植物は、光に依存しない酵素(光非依存型還元酵素)ももっているため、光がなくてもプロトクロロフィリドをクロロフィリドに変えることができてクロロフィル合成をつくることができるんです。つまり、frxC遺伝子を破壊してしまうと、光非依存型還元酵素がつくられなくなり、被子植物と同じ状況になってしまうことで、暗所でのクロロフィル合成ができなくなってしまったということです。
酸素をつくる光合成に酸素に弱い酵素が関わっているのはなぜ?
——ニトロゲナーゼと似ているのはなぜだろうという疑問から、ここまで研究が発展したのはすごいですね。
「被子植物は暗所で育てると緑色にならないのに、裸子植物ではきちんと緑色になるのはなぜか」という1世紀前の疑問がようやく解決できたことには、非常に感動しましたね。frxCの機能は、クロロフィルをつくる能力の違いを説明していたというわけです。frxCは、ゼニゴケにはあるのに、タバコ(被子植物)にはない遺伝子です。タバコはfrxC遺伝子を欠いているので、クロロフィルを暗所でつくる能力はありません。なお、frxC遺伝子はその機能にもとづき名前が変更され、現在はchlL遺伝子(またはbchL)とよばれています。
——この結果を基点にその後もずっと研究を進められたんですよね。
frxCがつくる光非依存型還元酵素を「DPOR(Dark-operative Pchlide oxidoreductase:DPOR)」と名付け、2010年には結晶構造解析を行い、その構造を明らかにしました。DPORの構造をニトロゲナーゼの構造と改めて比べてみると、やはりよく似ていますよね。
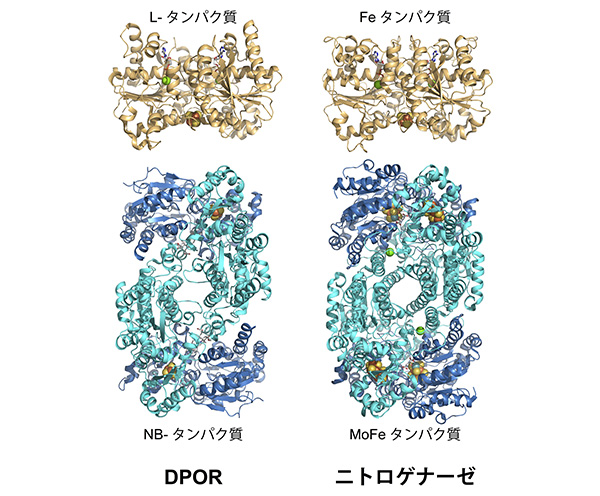
しかし、関わる反応はそれぞれ、光合成と窒素固定。プロトクロロフィリド分子内の二重結合を還元する反応と、窒素分子の三重結合を還元する反応ということで、まったく異なるものです。ただ、安定な化合物の多重結合を還元するという意味では似ているといえるかもしれませんね。
さらにDPORもニトロゲナーゼも、酸素に弱いというのが共通点です。でも、どうして酸素を発生する光合成生物の中で、酸素に弱いこれらの酵素は機能するのでしょうか? この矛盾を“酸素パラドクス”とよびます。私は、酸素をつくる光合成反応が酸素に弱い酵素に依存しているということに疑問をもちました。実は、窒素固定植物の研究をスタートしたのは、それが理由です。
ニトロゲナーゼも酸素に弱いはずなのに、光合成が起きているところで機能しているので、DPORと同じ課題を解決している酵素といえます。酸素に対する脆弱性と、酸素をつくる光合成との両立の仕組みを明らかにすることは、窒素固定能力を植物に付与することにつながるかもしれないなと思ったんです。
(後編へ続く)
空気を肥料にする「窒素固定作物」は、ハーバー・ボッシュ法を代替できるのか? – 名古屋大・藤田祐一教授インタビュー【後編】

名古屋大学大学院生命農学研究科 藤田祐一教授 プロフィール
1991年大阪大学大学院理学研究科博士課程修了。理学博士。大阪大学蛋白質研究所助手、米国インディアナ大学生物学科客員研究員、名古屋大学大学院生命農学研究科助教授、同准教授を経て、2016年より現職。2008年から2012年までJST戦略的創造研究推進事業さきがけ「光の利用と物質材料・生命機能」研究者(兼任)。
この記事を書いた人

- フリーランスライター/編集者。お茶の水女子大学大学院博士前期課程修了。修士(理学)。出版社でIT関連の書籍編集に携わった後、Webニュース媒体の編集記者として取材・執筆・編集業務に従事。2017年に独立。現在は、テクノロジー、ビジネス分野を中心に取材・執筆活動を行う。アカデミストでは、academist/academist Journalの運営や広報業務等をサポート。学生時代の専門は、計算化学、量子化学。 https://www.suto-hitomi.com/






