樹木と動物、菌類が歩んだ共生の歴史-大規模解析から見えてきた樹木の進化パターン-
花と果実の多様性
なぜ多くの植物は美しい花を咲かせ、美味しい果実を実らせるのでしょうか?
「虫を呼んで花粉を運んでもらい、動物たちに果実を食べて種子を運んでもらうため」――植物に詳しい方であれば、そんな答えが頭のなかに浮かぶかもしれません。
植物と花粉を運ぶ動物、または種子を運ぶ動物との関係性は、それぞれ送粉共生、種子散布共生と呼ばれます。植物が蜜や果肉などを動物に提供する代わりに、動物は花粉や種子の運搬を担います。実に地球上の9割以上もの植物が動物に依存して花粉を運搬しており、半数以上の樹種が動物に種子の運搬を頼っていると見積もられています。植物の花や果実の色や形態は種間で非常に多様であり、それらは共生している動物の形態や生態と密接に関係していることから、送粉共生や種子散布共生は植物の多様化の主要な原動力のひとつと考えられています。
このように多くの植物が花粉や種子の散布を動物に頼っており、植物の進化とも密接に関係している一方で、動物に依存することなく、風や重力に頼ることで、花粉や種子を散布する植物もいます。なぜ、送粉や種子散布を動物に頼る種と頼らない種がいるのでしょうか?また、このような送粉・種子散布様式の違いはそれぞれどのようなメリットがあるのでしょうか?

花粉と種子の散布距離
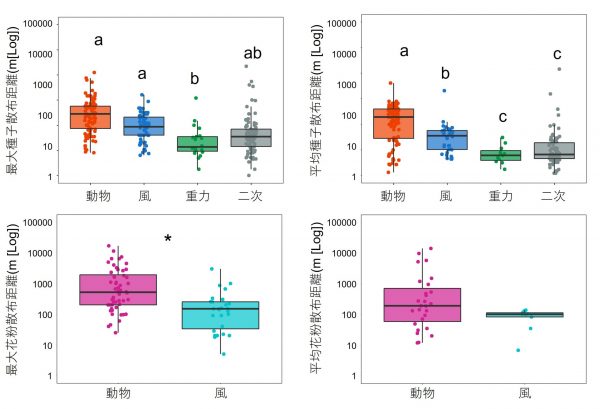
私たちは、動物による散布様式と風や重力による散布様式が生み出す違いについて、重要な要素として考えられている散布距離に着目しました。1970年から2020年までに出版された世界中の研究論文のなかから、樹木の花粉や種子の散布距離を直接測定している研究を収集し、1つひとつの論文を読み解き、樹種ごとの花粉と種子の散布様式(動物、風、または重力)と散布距離の情報を集め、散布様式のあいだで比較しました。
その結果、花粉と種子のどちらにおいても、動物による散布は風または重力による散布に比べて散布距離が長いという傾向が明らかになりました(図2)。つまり、樹木は送粉や種子散布を動物に依存することで、花粉や種子をより遠くへ散布することに成功しているようです。このように、動物との共生関係は、樹木にとって花粉や種子を遠くへ散布できるといったメリットがあることが分かってきました。
※ちなみにデータを解析する際に、一度地面に落下した種子を再び動物が散布する「二次散布」という現象についても散布距離を比較してみたところ、風や重力による種子散布と同程度であることが分かりました(図2)。ドングリなどの種子は、地面に落下した後にネズミやリスなどにより運ばれる(二次散布される)ことが良く知られていますが、遠くに種子が散布される効果よりも、より良い環境に散布されるなどの他のメリットの方が大きいのかもしれません。
このように散布距離という側面から多くの樹木が花粉や種子散布を動物に頼っている理由の1つが見えてきました。しかし、ここでさらなる疑問が浮かんできます。それは、「なぜ、すべての樹木が動物に花粉や種子の運搬を頼らず、一部の樹木は動物散布よりも散布距離が短い傾向にある風や重力による散布を行うのか?」ということです。たとえば、クルミの仲間やドングリを付けることで知られるブナ科の樹木は、花粉や種子の分散を風や重力に依存しています。動物に頼れば花粉や種子を遠くに散布できるにもかかわらず、なぜ、これらの樹木は、花粉や種子の散布を動物に頼らないのでしょうか?
私たちは、その理由として、植物にとって古くから重要な、もうひとつの共生関係である菌根共生が関与しているのではないかと考えました。
第三の共生関係:菌根共生の役割
植物が共生関係を結んでいるのは、実は昆虫や鳥などの動物だけではありません。菌根菌と呼ばれるカビやキノコの仲間は、植物から光合成で作られた糖分を受けとる代わりに、植物の根では直接届かない遠くの土壌や微細な土の隙間から水やリンなどの資源を吸収し、植物に提供しています。このような植物と菌根菌との共生関係を菌根共生と呼びます。この菌根共生もまた、送粉共生や種子散布共生と共にほとんどの植物種で見られる共生関係の1つです。菌根共生の歴史は古く、4億年以上昔に植物が上陸した直後から続いていることが知られています。
これまで、多くの研究者が植物の根を丹念に観察し、各植物種がどのような菌根菌と共生しているのかを記録してきました。その結果、樹木は種や属ごとに特定のタイプの菌根と共生していることが明らかになってきました。なかでも多くの樹木で観察されているのが、アーバスキュラー菌根(以下、AM)共生と外生菌根(以下、ECM)共生です。
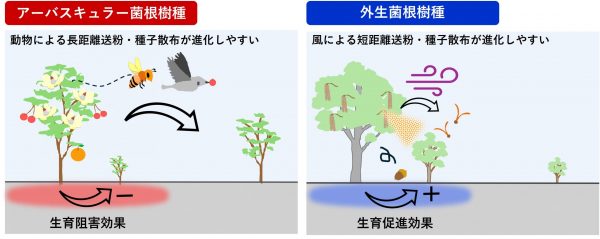
さらに、菌根共生タイプ(AM共生かECM共生か)によって、共生している樹木が周囲の同種他個体の生育に与える影響が異なることが指摘されています。AMと共生する樹種(以下、AM樹種)では、親株の周辺で病原菌や植食者が蔓延することにより、同種の芽生えの生育が阻害されやすく、一方、ECMと共生する樹種(以下、ECM樹種)では、親株周辺で同種の芽生えの生育が促進されやすいことが知られています。これは、ECMに樹木の芽生えを病原菌から効果的に保護する機能があるためだと考えられています。
私たちは、このような菌根共生がもつ同種に対する影響の違いが、送粉様式や種子散布様式の進化に強く影響しているのではないかと考えました。具体的には、AM樹種では、親株の周辺で同種の生育が阻害されるため、種子を親株から離れた位置に散布できる動物散布が進化しやすいと考えました。一方、親株近くで同種の生育が促進されるECM樹種では、散布距離が比較的短く、親株周辺に散布される風や重力による種子散布様式が進化しやすいと予測しました(図3)。
この予測を確かめるために、私たちは、ユーラシア、北米、南米、アフリカ、オセアニアなど世界各地に生育する樹木のうち、菌根共生のタイプ(AMまたは、ECM)が明らかになっている699種と、菌根共生のタイプが推定されている10,475種の樹木を対象に、送粉様式(動物媒、風媒)と種子散布様式(動物散布、風・重力散布)を花や果実の形態から推定しました。次に、菌根共生のタイプと送粉と種子散布様式の情報を系統樹上にマッピングし、対応関係を解析しました。その結果、私たちの予測したとおり、AM樹種は、動物媒花と動物による種子散布様式を示す種が、ECM樹種は、風媒花と風や重力による種子散布様式を示す種が多く、菌根共生のタイプと送粉・種子散布様式は互いに強く関連して進化していることが明らかになりました(図4)。
実際、日本の森林で菌根共生のタイプを意識しながら樹木を観察してみると、送粉様式、種子散布様式、菌根共生のタイプが関連していることを実感することができます(図5)。
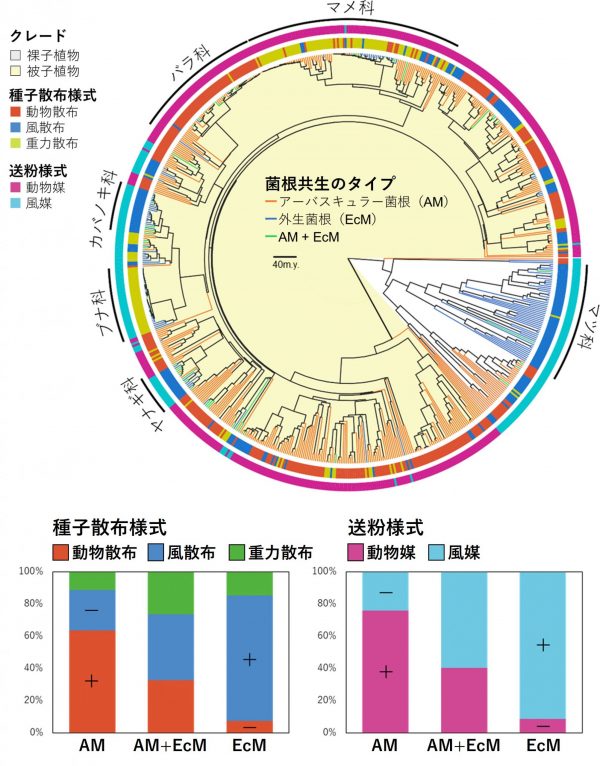

さらに私たちは、系統樹を用いて送粉様式、種子散布様式、菌根共生タイプの進化の因果関係についても解析しました。その結果、やはりAM樹種は、動物による送粉や種子散布を進化させやすく、ECM樹種は、風媒花と風や重力による種子散布様式を進化させやすいことが分かりました。
その一方で、私たちの予測に反して、送粉様式や種子散布様式もまた、菌根タイプの進化に影響していることが分かってきました。つまり、動物による送粉や種子散布様式を進化させた樹種は、AMとの共生を進化させやすく、風や重力による送粉や種子散布様式はECMとの共生を進化させやすいことが分かりました。このような送粉や種子散布様式の違いが菌根共生の進化に与える影響は、菌根菌のタイプ間での共生する樹木の特異性(共生できる植物の種の幅)の違いによって説明できるのではないかと考えています。AMは比較的幅広い植物分類群と共生関係を結ぶ一方で、ECMは特定の科や属を好む傾向が知られています。つまり、ECMでは親株の近くに種子が落下することで共生関係をより確実に構築できると考えられます。一方、動物によって親株から遠く離れた場所に種子が散布される樹木では、同種が生育しない地に散布されることも多いため、より広い植物分類群と共生関係を結ぶAMとの共生が適していたと考えることができます。
このように、菌根共生、種子散布共生、送粉共生は互いに影響を与え合いながら強く関連している様子が見えてきました。このような相互作用が、AM共生と動物による送粉・種子散布、ECM共生と風・重力による送粉・種子散布という、樹木の進化パターンを形成してきたと考えられます。
今回の発見は、植物の多様化や地球規模の生物多様性パターンの理解を大きく進展させる可能性を秘めています。これまでは、菌根共生、送粉共生、種子散布共生は、それぞれ独立に植物の多様化に寄与してきたと考えられてきました。しかし、実際には、これらの共生関係は互いに影響を与え合いながら、植物の多様化を促してきた可能性があり、植物の多様化プロセスを真に理解するためには、これら3つの共生関係の相互作用にも着目する必要があるのかもしれません。
また、菌根共生、送粉共生、種子散布共生には、熱帯から温帯にかけての緯度に沿ったパターンが存在することがそれぞれ異なる研究で報告されてきました。菌根共生においては、熱帯ではAM樹種が多く、温帯ではECM樹種が優占いるパターンが知られています。送粉や種子散布においては、熱帯では昆虫や鳥、哺乳類といった動物による送粉や種子散布を行う種が多く見られる一方で、温帯では動物に依存した送粉や種子散布が熱帯に比べて少ないというパターンが知られています。このような地球規模の生物多様性のパターンの形成にもまた、菌根共生、送粉共生、種子散布共生が互いに関わり合いながら進化してきたことが影響している可能性があります。
加えて、このような樹木の進化は、樹木の葉や幹などにおける性質の進化や、樹木の生育密度といった森林生態系のあり方、そしてそこに住む昆虫などのさまざまな生物の生態や進化にも影響を及ぼしてきたと考えられます。今後は、今回の研究で明らかになった樹木の進化パターンから、生態系の成り立ちに迫ることで、生物多様性のパターンやその形成プロセスの理解に貢献できるのではないかと考えています。
参考文献
- Yamawo A., Ohno M. (2024) Joint evolution of mutualistic interactions, pollination, seed dispersal mutualism, and mycorrhizal symbiosis in trees. New Phytologist, 243: 1290-1292.
この記事を書いた人

-
山尾 僚(やまお あきら)
京都大学 生態学研究センター 教授
2012年に鹿児島大学大学院連合農学研究科で植物の柔軟な防御応答に関する研究で博士号(農学)を取得。日本学術振興会特別研究員(PD)、弘前大学准教授を経て、現職につき、植物の進化や環境応答から生態系の仕組みを解き明かす研究を行っている。
HP:https://akira-yamawo.jimdofree.com/
大野 美涼(おおの みすず)
2024年、岩手大学大学院連合農学研究科にて樹木の冬季の日長認識に関する研究で博士号(農学)を取得。現在はPR会社に勤務。広報について現場で学びながら研究も継続して行っている。
HP:https://sites.google.com/view/misuzuohno/home
この投稿者の最近の記事
 研究成果2024年12月31日樹木と動物、菌類が歩んだ共生の歴史-大規模解析から見えてきた樹木の進化パターン-
研究成果2024年12月31日樹木と動物、菌類が歩んだ共生の歴史-大規模解析から見えてきた樹木の進化パターン-


