空気を肥料とする植物は可能か? – 光合成生物で窒素固定酵素を作動させる試み
ハーバー・ボッシュ法による窒素肥料生産と環境問題
窒素は、肥料の三大要素「窒素・リン酸・カリ」の筆頭にあげられ、植物の生産性を決める主な要因のひとつです。20世紀以降世界の人口は急速に増加し、現在75億に達しようとしています。この急速な人口増加は、1940年代から60年代にかけて収穫量の高いイネやコムギの栽培品種がつくられ、穀物生産量が飛躍的に増大したことで実現しました。これがいわゆる「緑の革命」です。これら栽培種に高い収穫量を発揮させるには、充分な窒素肥料が必須です。20世紀初頭にフリッツ・ハーバーとカール・ボッシュによって工業的窒素固定が確立され、この需要に応えられる窒素肥料供給が可能となっていたことが「緑の革命」の実現の必須要件でした。
ハーバー・ボッシュ法として知られる工業的窒素固定では、空気中の窒素を高温(500〜600℃)・高圧(200〜500気圧)条件、金属触媒存在下で水素と反応させてアンモニアに変換します。この反応条件の実現のために大量の化石燃料が消費されます。このため、工業的窒素固定により排出されるCO2量は莫大です。
作物が窒素肥料を最も必要とする時期に施肥のタイミングを合わせることは困難です。また、農家は充分な収穫を得るために過剰に化学肥料を与えてしまいがちです。このため、窒素肥料の大半は作物に吸収されず環境に流出してしまいます。窒素肥料成分は、環境で硝酸塩に変換され飲料水を汚染します。また、窒素不足に適応してきた生物群が、その生息環境に大量の窒素肥料成分が流入したために絶滅の危機にさらされています。また、窒素肥料成分は土壌細菌による脱窒作用の過程で窒素分子(N2)に変換されますが、この過程でCO2の300倍もの温室効果をもつ亜酸化窒素(N2O)が生じて大気に放出されます。このように、大量施肥による環境問題は看過できない状況にあります。
ハーバー・ボッシュ法が確立されて1世紀以上経過した今日、環境負荷が少なくより効率的な新しい窒素固定の方法が模索されています。
窒素固定を触媒する酵素ニトロゲナーゼを使った“窒素固定作物”作出への挑戦
大量のエネルギーを消費する工業的窒素固定に比べ、生物が行う窒素固定はニトロゲナーゼとよばれる酵素を使って行われ、常温常圧で進行します。ただし、窒素固定の能力をもつ生物は、原核生物と呼ばれる微生物の中の限られた種だけです。たとえば、根粒菌、土壌細菌アゾトバクター、シアノバクテリア、光合成細菌などが窒素固定菌として知られています。
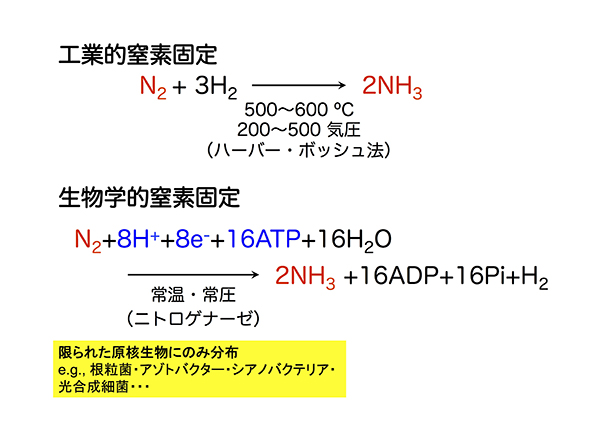
ハーバー・ボッシュ法に基づく工業的窒素固定では、高温高圧条件が必要とされるのに対し、ニトロゲナーゼによって触媒される生物学的窒素固定では、常温常圧で反応が進行する。
このような窒素固定菌がもつニトロゲナーゼを作物自身の細胞の中で作動させれば、まさに、空気中の窒素を自ら窒素肥料に変換して利用する“窒素固定作物”を作り出すことができるかもしれません。その最も直接的な方法は、ニトロゲナーゼの遺伝子を作物に導入して発現させることです。
しかし、その実現には大きな壁が立ちはだかっています。まず、ニトロゲナーゼは、酸素によって急速に壊されてしまうという性質を持っています。作物は光合成生物ですから、光合成で酸素を作ります。そのような作物の細胞の中ではニトロゲナーゼはすぐに壊されてしまいます。
さらに、ニトロゲナーゼは、その分子の内部に鉄モリブデン補因子(FeMo-co)と呼ばれる鉄・硫黄・モリブデン・炭素・有機酸から成る複雑な金属触媒をもっており、まさにその補因子の上で窒素がアンモニアに変換される反応が起こります。つまり、FeMo-coがなければニトロゲナーゼは活性が発揮できないただのタンパク質の塊に過ぎません。
問題は、FeMo-coをつくるためには多数のタンパク質の作用が必要とされる点にあります。ニトロゲナーゼ自身は、3つのタンパク質の複合体ですから遺伝子は3つですが、FeMo-coをつくるためにさらに8個以上のタンパク質が必要とされます。つまり、窒素固定性作物を作り出すためには、酸素に対する脆弱性、多数の遺伝子の必要性、という2つの大きな壁を打ち破る必要があります。
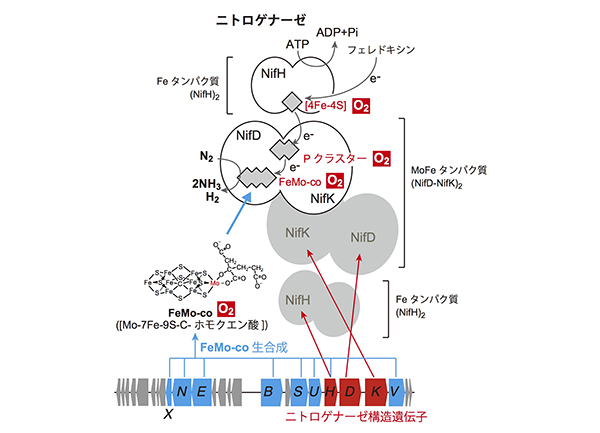
ニトロゲナーゼは、Feタンパク質(NifH二量体)とMoFeタンパク質(NifD-NifKヘテロ4量体)という2つのタンパク質成分から構成され、反応に必要とされる電子は、3つの金属クラスター([4Fe-4S]、Pクラスター、FeMo-co)を介して伝達され、最終的にFeMo-co上で窒素分子の還元反応が進行する。これらの金属クラスターはすべて酸素によって速やかに破壊されてしまう(赤い背景の“O2”で示す)。また、ニトロゲナーゼの3つの構造遺伝子に加え、FeMo-coの生合成に少なくとも8つの遺伝子が必要とされる。
モデル実験としてのシアノバクテリアへの遺伝子導入
シアノバクテリアは、植物と同じく酸素を作る光合成を行う原核生物です。しかも、シアノバクテリアの約半数種は窒素固定の能力をもっています。つまり、窒素固定性シアノバクテリアは、光合成と窒素固定を両立させている唯一の生物群です。また、シアノバクテリアは、植物の葉緑体の起源となった生物と考えられています。したがって、シアノバクテリアで得られた知見は、葉緑体でニトロゲナーゼを発現させるために大いに役に立つに違いありません。そこで、私たちは、シアノバクテリアを使ったモデル実験を考案しました。窒素固定性シアノバクテリアのニトロゲナーゼ遺伝子を、もともと窒素固定の能力をもたないシアノバクテリアに導入することで窒素固定能を付与する、という実験です。
私たちは、この実験に先立ち、プレクトネマという窒素固定性シアノバクテリアを使って窒素固定遺伝子の発現制御について研究をしてきました。その過程で、プレクトネマのゲノムに窒素固定に関わる遺伝子が集積した窒素固定遺伝子クラスターを見つけ、さらにCnfRという転写制御タンパク質も発見しました。これらの遺伝子を、窒素固定の能力をもたないシアノバクテリア(シネコシスティス6803)にまとめて導入して、窒素固定能を付与しようというアイディアです。
ついにニトロゲナーゼ活性を検出
プレクトネマの窒素固定遺伝子クラスターは約20kbにわたっており、この長さのDNAを一度にシネコシスティス6803に導入することは困難です。そこで、5本の断片に分けて1本ずつ順番に導入するという地道な実験を進め、5段階でようやく20.8kbのDNA断片(24個の遺伝子が含まれます)全体の導入を完了しました。最後に、転写制御タンパク質CnfRの遺伝子を導入し、形質転換体CN1を得ました。さらに、1個もしくは4個の遺伝子を加えた各々CN2とCN3も作りました。これらの形質転換体のニトロゲナーゼ活性(アセチレン還元活性)を測定しましたが、当初はまったく活性が見られず、落胆しました。しかし、脱酸素試薬(ジチオナイト)の添加や反応時間の延長など、測定条件を見直すことで、ついに有意な活性を検出することに成功しました。
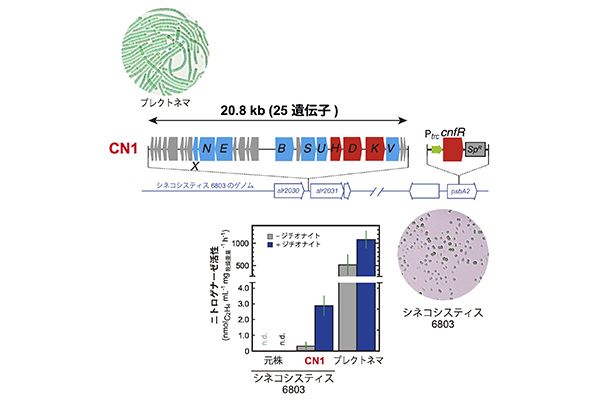
プレクトネマの20.8kbにわたる窒素固定遺伝子クラスターとその制御タンパク質CnfRをコードする遺伝子を導入したシネコシスティス6803の形質転換体CN1で、低いが有意なニトロゲナーゼ活性が検出された
活性は3つの形質転換体のすべてで見られましたが、CN1が一番高い活性を示しました。ただ、最も高い活性でもプレクトネマの約0.3%(乾物重量当たり)という低い値にとどまっています。低い活性のせいか、残念ながらいずれの形質転換体も窒素固定的条件では生育することができません。
これからの展望
今後は、シネコシスティス6803の形質転換体(CN1〜CN3)の活性をより向上させて、窒素固定的生育を実現することを目指します。その過程で、窒素固定能を付与するために必要とされる遺伝子セットが明らかになります。その知見を元にして、次は、緑藻やモデル植物(クラミドモナスやシロイヌナズナなど)に遺伝子を導入し、窒素固定能をもった藻類や植物の作出を実現させたいと考えています。そうなれば夢の“窒素固定作物”が現実となる日も近いはずです。
参考文献
- Tsujimoto, R., Kamiya, N., and Fujita, Y. (2014) Transcriptional regulators ChlR and CnfR are essential for diazotrophic growth of nonheterocystous cyanobacteria. Proc. Natl. Acad. Sci. USA 111, 6762-6767.
- Good, A. (2018) Toward nitrogen-fixing plants: A concerted research effort could yield engineered plants that can fix nitrogen. Science 359, 869-870.
- Tsujimoto, R., Kotani, H., Yokomizo, K., Yamakawa, H., Nonaka, A. and Fujita, Y. (2018) Functional expression of an oxygen labile nitrogenase in an oxygenic photosynthetic organism. Sci. Rep., 8, 7380, DOI: 10.1038/s41598-018-25396-7
この記事を書いた人

-
名古屋大学大学院生命農学研究科・教授。
大学院生の頃に、ニトロゲナーゼと類似したタンパク質の葉緑体での機能を明らかにしようとする研究を始めました。その際、葉緑体のモデルとしてシアノバクテリア(プレクトネマ)を使い始めました。このニトロゲナーゼ類似タンパク質がクロロフィル生合成に関わる酵素であることを明らかにしたことで、研究はクロロフィル生合成に展開していきました。その過程で、プレクトネマの形質転換系やゲノム決定などの技術的環境の整備も行ってきました。最近、これまで行ってきた研究が、窒素固定という形で収斂しつつ発展しているように思え、研究のおもしろさ・奥深さを実感しています。



