チンパンジーの細胞をリプログラミング – iPS細胞製作の副産物が示す神経堤細胞様の特性
出発点:ヒトの特性を理解するためには何が必要だろうか?
筆者(今村)が博士号を取得して数年が経ったころです。それまでマウスを扱った基礎医学研究に取り組んできましたが、新たにコモンマーモセットという小型のサルを扱い始めたことを機に、次の疑念をもつようになりました。
ひとつは、マウスと霊長類では思った以上に違いが大きいことが段々と見えてきて、「マウス生物学」に取り組んでいても結局ヒトのことはわからないのではないか、ということ。至極当たり前の懸念ではあるのですが、長らくマウスの研究ばかりをしていた結果、「マウスの研究さえすれば、ヒトや哺乳類のこともわかる」と半ば信じ込んでいました。
2つ目は、ヒトだけを見ていてもヒトのことはわからないのではないか、という疑問です。特に疾病の研究では、健常と罹患の比較によって病態のメカニズムを探ろうとしますが、決定的な原因遺伝子などの病気の本質を捉えることは非常に困難です。もしかすると、ヒトの健常と罹患のあいだの違いはそれほど大きくないのではないか。だとすれば、病気の本質がなかなか捉えられないのは、対照が適切ではないことに理由があるのではないか。そのようなことを考えていました。
そうした折、当時大学院生だった林煜欽くんが日本分子生物学会のブース展示にて、京都大学霊長類研究所の共同利用研究制度を使って霊長類サンプルが利用できる、との情報を入手してきました。
そうか、チンパンジーという手があるのか。ヒトにもっとも近縁な現生動物であり、およそ99%のゲノム情報を共有するチンパンジー。この外群をとおしてヒトをみれば、ヒトの特性の本質が見えてくるかもしれない。それまで考えもしなかった可能性でしたが、チャレンジし甲斐のある課題だと感じました。
チンパンジー培養細胞をリプログラミングしてみた
さっそく、霊長類研究所の共同利用研究に申請し、国内の動物園で亡くなったチンパンジーの組織試料を使わせていただきました。最初の目的は、チンパンジーiPS細胞の作製です。チンパンジーiPS細胞を作製すれば、発生・分化を再現してヒトと比較することができます。
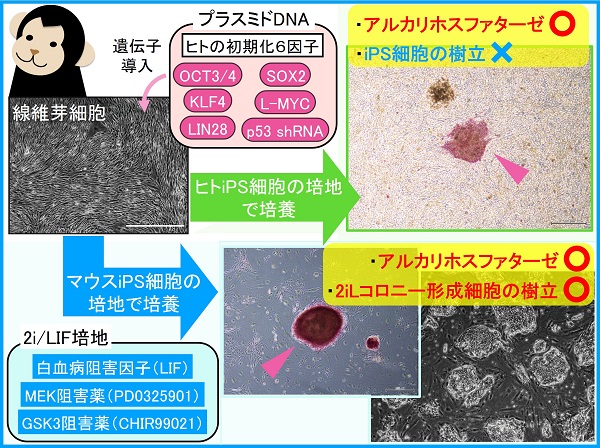
まずは、チンパンジーの培養細胞に蛍光タンパク質のプラスミドを入れ、問題なく遺伝子導入ができることを確認しました。次に初期化因子のプラスミドを導入してみました。しかし、なぜかiPS細胞が樹立できません。多能性幹細胞の指標のひとつであるアルカリホスファターゼを染色してみると、果たして陽性の細胞集団が検出されるので、リプログラミングはされているようでした。はて、どうしたものか。
ひとつの試行として、マウスのiPS細胞を培養する条件も試してみました。後述するように、マウスとヒトではiPS細胞の多能性の状態が違い、そのため培養液も違います。おまけ程度のつもりで、マウスiPS細胞用の通称2i/LIF培地(白血病阻害因子(LIF)、MEK阻害薬(PD0325901)、GSK3阻害薬(CHIR99021)を添加した化学合成培養液)で培養してみたところ、マウスiPS細胞と似たドーム状のコロニーを作る細胞を得ることができました。以降、この細胞を「2iLコロニー形成細胞」と呼ぶことにします。
ナイーブ型とプライム型:多能性幹細胞が示す分化多能性の2つの状態
ここで、多能性幹細胞(ES細胞やiPS細胞など)が示す分化多能性について簡単に説明します。哺乳類の分化多能性は2つの状態に大別され、ナイーブ型、プライム型と呼ばれています。
カタカナで書くと意味が掴みにくいのですが、ナイーブはnaïve、プライムはprimedが語源で、分化の方向に「感作されていない(ナイーブ)」あるいは「感作された(プライム)」状態を表しています。したがって、ナイーブからプライムへと発生段階が進行し、ナイーブ型は着床前、プライム型は着床後の胚を反映します。また、両者の性質には分子・細胞レベルでさまざまな違いがあります。
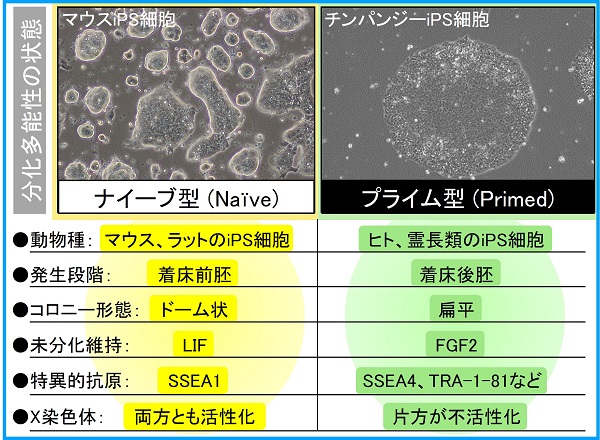
例を挙げると、ナイーブ型の細胞はドーム状のコロニーを形成するのに対し、プライム型では扁平なコロニーを作ります。また、ナイーブ型の未分化状態はLIFシグナルによって維持されますが、プライム型では線維芽細胞増殖因子2(FGF2)シグナルに依存します。他にも、雌性ゲノム(XX)をもつ場合、ナイーブ型では両方のX染色体が活性化している一方、プライム型では体細胞と同じく片方が不活性化している、などの違いがあります。
通常、マウスのiPS細胞はナイーブ型、ヒトや霊長類のiPS細胞はプライム型の特徴を示します。マウスでは培養条件を切り替えるだけでナイーブ型とプライム型を容易に転換することができますが、ヒトのナイーブ型を誘導することは非常に困難で、複数の遺伝子導入と薬剤処理を組み合わせることが必要です。ヒトiPS細胞をマウスiPS細胞(ナイーブ型)用の2i/LIF培地で培養した場合、初期神経上皮細胞のような性質をもった細胞に変化してしまいます。
では、2i/LIF培地でリプログラミングしたチンパンジーの2iLコロニー形成細胞は、一体どのような性質をもっているのでしょうか?
2iLコロニー形成細胞は、多分化能・腫瘍形成能・神経堤細胞様の性質をもつ
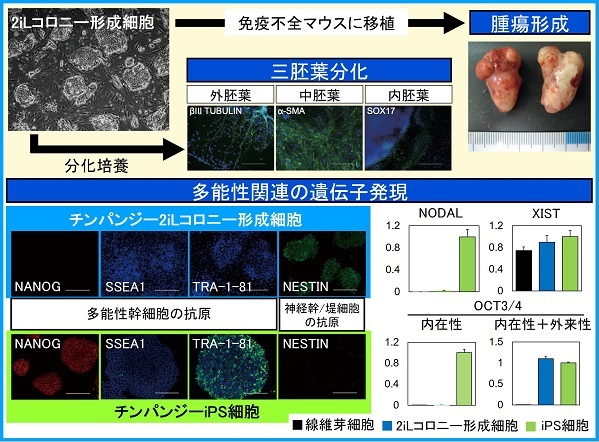
チンパンジーの2iLコロニー形成細胞は、ナイーブ型のマウスiPS細胞と同じような感覚で培養することができ、正常な染色体を維持して長期間増殖します。また、胚様体形成による分化培養によって三胚葉(外胚葉、中胚葉、内胚葉)の細胞に分化し、免疫不全マウスに移植すると三胚葉性の腫瘍を形成します。
ところが、遺伝子発現を解析してみると、この細胞は多能性幹細胞の抗原であるNANOGや細胞膜表面糖鎖(SSEA1、TRA-1-81など)が陰性であると同時に、神経幹細胞や神経堤細胞の抗原であるNESTINが陽性であることがわかりました。
さらに詳しく解析すると、多能性関連遺伝子(NODAL、REX1など)の発現があまり見られないこと、X染色体の不活性化を引き起こすXISTが発現していることがわかりました。一方、初期化因子として導入した遺伝子では、OCT3/4の外来遺伝子がゲノムに挿入されており、内在性遺伝子の代わりに発現していました。
体細胞がiPS細胞へとリプログラミングされる際、間葉上皮転換という現象が起こります。このとき、間葉系の遺伝子の発現が抑えられ、代わりに上皮系の遺伝子の発現が誘発されます。しかし、2iLコロニー形成細胞では間葉系遺伝子が一部発現しており、また上皮系遺伝子の発現が誘導されていない状態にありました。
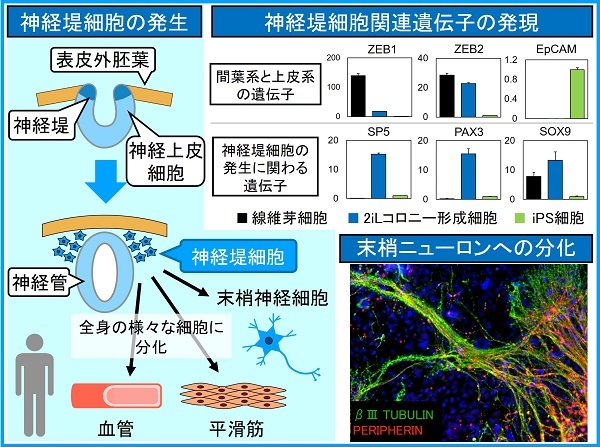
間葉系の遺伝子発現は、神経堤細胞の特徴でもあります。神経堤細胞とは、胎児の神経管が形成される時期に神経板と表皮外胚葉の間から生じる細胞で、全身のさまざまな細胞(顔面、血管、平滑筋、末梢神経系など)に分化することから「第4の胚葉」とも呼ばれます。
予想外なことに、2iLコロニー形成細胞では神経堤細胞の発生に関わる一連の遺伝子が発現していました。さらに、神経系への特異的な分化誘導を行ったところ、iPS細胞では脳のニューロンが分化するのに対し、2iLコロニー形成細胞では神経堤細胞から派生する末梢ニューロンへと分化することがわかりました。
結語:リプログラミング機構の解明に向けて
今回の研究では、2i/LIF培地でチンパンジーの細胞をリプログラミングすると、多分化能と腫瘍形成能をもつ細胞が誘導されるものの、この細胞は多能性幹細胞ではなく、神経堤細胞様の特徴を併せもつ細胞であることがわかりました。
この細胞は、末梢ニューロン研究のツールとして有用であると考えられます。では、なぜ神経堤細胞の特性をもつのでしょうか? 可能性としては、iPS細胞に至るリプログラミングの中間段階と、iPS細胞とは別方向へのリプログラミングの2つが考えられます。
iPS細胞にリプログラミングされるとき、原条という発生状態を経ることがわかっています。また、iPS細胞から原条を分化誘導する場合、NODALとWNTの2つのシグナルの活性化が必要で、NODALシグナルが阻害されると神経堤細胞の方向に分化します。
今回、2i/LIF培地によってWNTシグナルが活性化される一方で、NODALの発現は誘導されていないことから、リプログラミングの途中で原条から神経堤細胞の方向に脱線したのかもしれません。しかし、ヒトiPS細胞を2i/LIF培地で培養した過去の研究では、神経堤細胞ではなく初期神経上皮細胞様の特性を示しています。これについては、初期化因子やベクターの種類、種差などの違いの影響も考えられます。このように残された謎は多く、リプログラミング機構の理解に向けてさらなる研究が必要です。
ちなみに、本来の目的だったチンパンジーiPS細胞の作製も成功しており、ヒト特性の解明に向けた研究が進んでいます。それらの研究成果については、また別の機会に。
参考文献
- Zachary Yu-Ching Lin, Risako Nakai, Hirohisa Hirai, Daiki Kozuka, Seiya Katayama, Shin-ichiro Nakamura, Sawako Okada, Ryunosuke Kitajima, Hiroo Imai, Hideyuki Okano, Masanori Imamura (2020). Reprogramming of chimpanzee fibroblasts into a multipotent cancerous but not fully pluripotent state by transducing iPSC factors in 2i/LIF culture. Differentiation, 112: 67-76.
- 今村公紀、仲井理沙子『ニホンザルのiPS細胞の作製に成功!-「霊長類学」の新たな可能性』(academist journal、2018年10月23日)
この記事を書いた人

-
今村公紀(写真左)
京都大学ヒト行動進化研究センター 助教、博士(医学)。富山県高岡市出身。金沢大学理学部、奈良先端科学技術大学院大学、京都大学大学院医学研究科、三菱化学生命科学研究所にて学生時代を過ごした後、滋賀医科大学 特任助教、慶應義塾大学医学部 特任助教、理化学研究所 客員研究員、京都大学霊長類研究所 助教を経て、2022年より現職。
仲井理沙子(写真右)
理化学研究所バイオリソース研究センターiPS創薬基盤開発チーム 特別研究員、博士(理学)。京都府舞鶴市出身。富山大学理学部卒業。日本学術振興会特別研究員(DC2)を経て、京都大学大学院理学研究科修了。2022年より現職。
研究室ウェブサイト





