【特別寄稿】深海生物「テヅルモヅル」とは何者か?
これまでacademist journalにテヅルモヅルに関する記事をいくつか寄稿してきたが、せいぜい「深海生物である」という説明にとどまっていたように思う。本稿では、テヅルモヅルとはどういう生きものであるかを、学術的にみなさまにご説明したい。
テヅルモヅルの分類
テヅルモヅルは、学術上、「棘皮動物門(もん)のクモヒトデ綱(こう)」に分類される。棘皮動物とは、ウニ、ナマコ、ヒトデを含むグループであり、クモヒトデとは、腕が細長いヒトデのような形をした生きものである。
その形や、クモ「ヒトデ」という名前のせいか、よく「クモヒトデはヒトデのいちグループですか?」と聞かれることがあるが、ヒトデとクモヒトデは「綱(門のひとつ下の階級)」のレベルで分けられる別の分類群である。我々に近いところに置き換えてみると、魚綱と両生綱くらいの違いに相当する(2)。
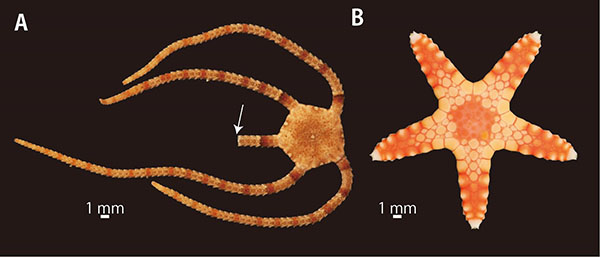
星形で、一見するとよく似ているヒトデ綱とクモヒトデ綱だが、実は腕の構造がまったく異なる。海で星形の生物を発見したら、ひっくり返して口側をよく観察してほしい。星形の真んなかに口があるが、そこから腕の正中線上に伸びる溝があればヒトデ、なければクモヒトデである。
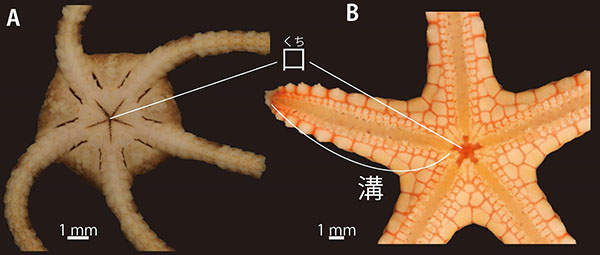
非常に簡単な違いに思えるが、これは、腕の内部の骨格の構造を反映した本質的な違いである(6)。他にも、顕微鏡での観察を要することが多いが、クモヒトデには肛門がなく、ヒトデにはある(肛門がないヒトデもいるが)。多孔体と呼ばれる、海水を体内の水管と呼ばれる管に取り込む入口の骨片板が、クモヒトデでは口側に、ヒトデでは反口側にある、といった違いもある。テヅルモヅルは、腕の口側に溝が無く、肛門を欠き、多孔体が口側にあるので、立派なクモヒトデなのである。ちなみに、クモヒトデは英語では”Brittle star”、和訳すると「脆いヒトデ」と呼ばれている。これは、クモヒトデが刺激に反応して腕を簡単に自切する様子にちなんでいる。
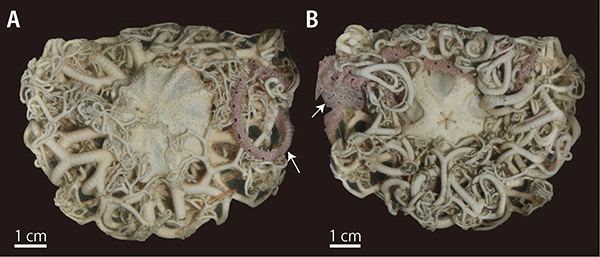
テヅルモヅルは、クモヒトデ綱のなかのツルクモヒトデ目(もく、綱のひとつ下の階級)に含まれる。他の目との関係についてはここでは詳しくは語らないが、ツルクモヒトデ目は他の目とは形態的に、「腕針(わんしん)」の配置で見分けることができる。
腕針とは、クモヒトデの腕に生えている微小な骨片で、ウニの棘が小さくなったようなものや鉤爪状など、形はさまざまである。この腕針の生じる位置が、ツルクモヒトデ目では腕の口側なのに対し、その他のクモヒトデでは腕の側面なのである(7)。ツルクモヒトデ目は、その名のとおり「蔓(つる)」のように腕をサンゴなどに巻き付けて暮らしており、この腕針の配置は、サンゴに絡みやすいように進化したものなのかもしれない。ちなみに、目レベルの違いとは、哺乳類に当てはめてみると、ウサギ目とネズミ目くらいの違いということである(2)。
テヅルモヅルとは、ツルクモヒトデ目のなかの腕が分岐するクモヒトデである。何とも珍妙な形ではあるが、その腕を網状に海中に広げることで、効率よくプランクトンなどを捕らえて食べていると考えられている。

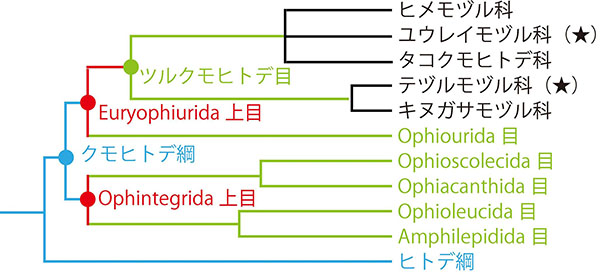
しかし、テヅルモヅルは、進化的にはまとまった分類群ではない。ツルクモヒトデ目は現在5つの科(か、目のひとつ下の階級)に分けられているが(8)、テヅルモヅルはそのうち、系統的に遠縁な2つの科に別々に存在するのである。ツルクモヒトデ目の祖先は、腕が分岐していなかったと考えられているため(5)、そのような祖先から、少なくとも2回、「テヅルモヅル化」が起こったものと考えられる。ちなみに、科レベルの違いといえば、哺乳類でいえば、イヌ科、クマ科、アシカ科くらいの違いに相当する(2)。
テヅルモヅルの生態
紅海でサメハダテヅルモヅル(Astroboa nuda)を観察した研究例によれば、彼らは昼間には特定の岩の隙間などに腕を丸めて潜んでおり、日が暮れるとそこから抜け出し、潮の流れの早い場所(岩やサンゴのてっぺんなど)に移動し、夜間ずっと、その腕を海中に広げて餌をとり続けるらしい。そして朝日と共に、また元いた場所に帰っていくということだ(3)。ちなみに何を食べているかは詳しくはわかっていないが、水族館ではオキアミなどを餌としてあげているらしい。ツルクモヒトデ目は190種近くが知られているが、詳しい経時的な観察例があるのはこのAstroboa nudaや、大西洋産のGorgonocephalus captmedusaeくらいである(10)。その他、腕の動かし方や(1)、発生様式の一部について(9)、ごく限られた種に関する研究がある以外、ほとんどが謎に包まれたままになっている。
なぜこれほどまでに研究が進んでいないのか?
私自身、テヅルモヅル類の研究を始めてみて、その理由を身を持って知ることとなった。第一の理由として、深海性の種が多いことがあげられる。浅瀬でも見られる種はいるが、磯を歩いていて見つけられることはほとんどなく、少なくともスキューバダイビングでないと彼らの元へは到達できない。しかも上記したとおり多くが夜行性のため、昼間に見つけても岩の奥深くに潜んでしまっているので引っ張り出すのは難しく、腕を広げる勇猛な姿には滅多にお目にかかれない。このようなテヅルモヅル類を収集するため、筆者は何度もスキューバダイビングを行ったり、乗船調査に参加してきたが、採集できた例はあまり多くない。20日間船に乗ってボウズということもあった。
第二の理由として、飼育が難しいことがあげられる。流れが速い場所に生息するテヅルモヅルはキレイな水を好むらしく、彼らが好むような海流や水質を飼育条件下で再現するのは、水族館でもなかなか難しいらしい。生殖発生や行動などの飼育がほぼ不可欠な研究は、このような理由で進んでいない。
最後の理由は、彼らが大型であるということである。腕を広げると1m以上になる種もザラにいるため、採集した標本用の容器や、保存用のエタノールの量、そしてそれらを確保しておくスペースの確保がバカにならない。筆者の研究室では大型の密閉性の高いバケツを使ったりしているが、10数リットルのエタノールが入ったバケツは相当な重さであり、観察のたびにそれらのバケツの運搬に結構な労力を払うことになる。このような、他の小型動物にはない苦労を強いられるところも、テヅルモヅル類研究が敬遠される理由のひとつであろう。
しかし、苦労はあるが、「謎=学術的課題」であるわけで、テヅルモヅルこそ研究成果の宝庫といえる。実際、テヅルモヅルの秘密が明らかにできた際の喜びは筆舌に尽くしがたいものがある。この珍妙な動物については、拙著『深海生物テヅルモヅルの謎を追え!―系統分類から進化を探る―』(東海大学出版会)に、これまでの研究史の一部を紹介している。テヅルモヅルにご興味を持たれた方は、参考にしていただければ幸いである。また、テヅルモヅルの研究の現状については、オンラインで筆者による解説がなされている。英語で、購読が必要ではあるが、さらに深く学んでみたい方はそちらも参考にしていただきたい。
引用文献
1. Hendler, G. (1982) Slow flicks show star tricks: Elapsed-time Analysis of bastketstar (Astrophyton muricatum) feeding behavior. Bulletin of Marine Science. 32 (4): 909–918.
2. 増田隆一. (2017) 哺乳類の生物地理学. 東京大学出版会. 183 pp.
3. Tsurnamal, M. & Marder, J. (1966) Observations on the basket star Astroboa nuda (Lyman) on coral reefs at flat (Gulf of Aqaba). Israel Journal of Zoology 15:9–17.
4. O’Hara, T. D., Hugall, A. F., Thuy, B., Stöhr, S. & Martynov, A. V. (2017) Reconstructing higher taxonomy using broad-scale phylogenomics: The living Ophiuroidea. Molecular Phylogenetics and Evolution. 107: 415–430.
5. 岡西政典. (2016) 深海生物テヅルモヅルの謎を追え!-系統分類から進化を探る-. フィールドの生物学シリーズ20. 東海大学出版会. 299 pp.
6. 岡西政典. (2016) 分子系統と形態観察から探る深海性クモヒトデ類(棘皮動物門)の進化. 化石研究会会誌. 49 (1): 26–34.
7. Okanishi, M., O’Hara, T. & Fujita, T. (2011) Molecular phylogeny of the order Euryalida (Echinodermata: Ophiuroidea), based on mitochondrial and nuclear ribosomal genes. Molecular Phylogenetics and Evolution. 61: 392–399.
8. Okanishi, M. & Fujita, T. (2013) Molecular phylogeny based on increased number of species and genes revealed more robust family-level systematics of the order Euryalida (Echinodermata: Ophiuroidea). Molecular Phylogenetics and Evolution. 69: 566–580.
9. Patent, D. H. (1970) Life history of the basket star, Gorgonocephalus eucnemis (Müller & Troschel) (Echinodermata; Ophiuroidea). Ophelia. 8 (1): 145-159
10. Rosenberg, R., Dupont, R., Lundälv, T., Sköld, H.N., Norkko, A., Roth, J., Stach, T. & Thorndyke, M. (2005) Biology of the basket star Gorgonocephalus caputmedusae (L.). Marine Biology. 148: 43–50.
この記事を書いた人
- 茨城大学理学部助教. 理学博士. 1983年生まれ. 東京大学大大学院理学系研究科生物科学専攻博士課程修了. 文部科学省教育関係共同利用拠点事業研究員(京都大学瀬戸臨海実験所)を経て, 現職.
この投稿者の最近の記事
 研究成果2017年7月28日【特別寄稿】深海生物「テヅルモヅル」とは何者か?
研究成果2017年7月28日【特別寄稿】深海生物「テヅルモヅル」とは何者か? 研究成果2017年2月6日和歌山県・白浜沖に秘められた生物多様性 – 未知の種が10種以上見つかる
研究成果2017年2月6日和歌山県・白浜沖に秘められた生物多様性 – 未知の種が10種以上見つかる お知らせ2016年6月24日キヌガサモヅルの分類はどこまで進んだのか? – academistの第一弾プロジェクトは今
お知らせ2016年6月24日キヌガサモヅルの分類はどこまで進んだのか? – academistの第一弾プロジェクトは今


