精子はどのように卵を見つけるか? – ホヤが受精するときの繊細な認識機構
精子の走化性
ヒトをはじめ多くの動物は性を持ち、雄由来の精子と雌由来の卵とが受精という融合をおこすことによって新しい世代を生み出します。その過程で精子は卵を探す必要がありますが、精子は雄の体外に出るとわずかな時間しか運動することができません。
そこで精子が卵に無事たどり着けるように、卵または雌性器官が自身の場所を示す化学物質を放出し、そのシグナルを手がかりに精子が卵まで誘導される現象が見られます。これは精子走化性と呼ばれ、植物から動物まで広く見られる現象で、特に海水中に放卵・放精を行って体外受精を行う海産動物で顕著に見られます。また、多くの動物において精子の誘引作用は種特異的であり、精子走化性は、精子が同種の卵と出会う確率を上げる仕組みと考えられています。
精子誘引物質と精子の運動
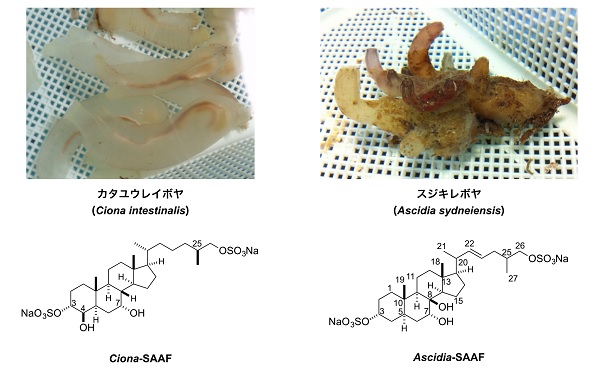
走化性が多くの動物で知られている一方、卵から放出される精子誘引物質はごく微量であるため、その同定は難しく、精子誘引物質が同定されている動物は、ホヤ、ウニ、サンゴ、アワビ、ヒトデなど、まだ10種程度しかありません。また同定された精子誘引物質は、タンパク質性の大きな分子から低分子有機化合物まで多様であることもあり、精子走化性の分子機構はまだあまりわかっていません。そのなかで、研究が良く進んでいる動物が、棘皮動物であるウニの一種 (Arbacia puncturata)(大西洋産のウニ)と、尾索動物であるカタユウレイボヤ(Ciona intestinalis)です。
走化性運動時の精子はらせん運動をしながら段々と卵へ近づいていきますが、その際、誘引源から遠ざかると遊泳方向が急激に変化し、引き続いて誘引源方向へと直進するという特徴的な方向転換(ターン)を行います。この「ターン」の際に、精子鞭毛内でCa2+の一過的な上昇(Ca2+バースト)がおこり、それが鞭毛波形を瞬時に変化させ、遊泳方向の転換を行っていることがウニやホヤでわかっています。
またウニにおける精子誘引物質は卵の周りにあるゼリー層に含まれるペプチドであり、これによって精子が誘引されることが知られています。このペプチドは、精子の受容体であるグアニル酸シクラーゼ(cGMPの合成酵素)を活性化してcGMPを作り出し、cGMPは最終的にCa2+チャネルのCatSperを開口させることがこれまでに示されています。
一方、ホヤの精子誘引物質は、カタユウレイボヤおよびスジキレボヤ(Ascidia sydneiensis)において「SAAF(Sperm-Activating and -Attracting Factor)」と呼ばれるステロイド誘導体であることがわかっています。
ホヤでは精子の受容体は不明でしたが、グアニル酸シクラーゼが精子走化性に関与していることは否定的に考えられています。したがって、ホヤを含め、ウニ以外の動物ではどのように精子誘引物質がCa2+濃度を調節しているのかは長年の謎でした。さらに、精子誘引物質がCa2+チャネルの開口につながるとすると、なぜ精子が卵から遠ざかる際にCa2+バーストを引き起こすのか、その仕組みも説明がつきませんでした。
ホヤの精子誘引物質はCa2+ポンプに作用する
この疑問を解消するため、我々は、カタユウレイボヤにおいて精子誘引物質SAAFがどのように精子に働きかけるかの解明を進めてきました。そして、カタユウレイボヤではSAAFは精子膜に存在するCa2+ポンプである「PMCA(Plasma Membrane Ca2+-ATPase)」と特異的に結合することが明らかとなりました。
PMCAは精子鞭毛に多量に存在し、SAAFと結合することで酵素活性が上昇しました。さらに、PMCAの阻害剤を精子に作用させると、SAAFに対する走化性を示さなくなること、その際には精子内のCa2+濃度が高いままに保たれていることがわかりました。
これらのことから、カタユウレイボヤの精子誘引物質は、精子細胞膜にあるCa2+ポンプ(PMCA)に作用し、精子内のCa2+濃度を調節していることが明らかとなりました。
精子誘引物質SAAFは、精子に作用しているあいだはPMCAを活性化して精子内のCa2+濃度を下げる方向に働いており、精子が誘引源より遠ざかり、誘引物質の濃度が減少するとPMCAの不活性化によりCa2+排出が止まり、結果的にCa2+バーストにつながることを示しています。このような繊細な制御は、SAAFとPMCAの“弱い”結合によってもたらされると考えられます。
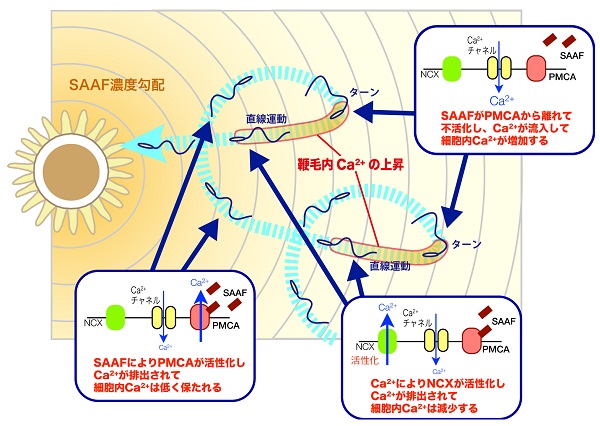
従来、細胞膜のCa2+ポンプにはCa2+流入で起こった情報伝達を終了させるために、細胞外へCa2+を排出する役割があることしかわかっておらず、積極的にシグナルを生成していることが示されたのは初めてです。
今後、精子誘引物質とCa2+ポンプの相互作用の種特異性や、Ca2+の詳細な調節機構を明らかにすることによって、卵に対する精子走化性の分子メカニズムの詳細とその進化の理解を進めると同時に、細胞内Ca2+濃度を外部から制御するための新たな手法の開発が期待されます。
受精における種特異性の分子機構解明に向けて
本研究により、精子走化性において誘引物質が精子内Ca2+濃度を調節するしくみの一端が明らかとなりました。また、誘引物質がCa2+濃度の減少を誘導することで、精子が誘引物質の減少という負の濃度変化の感知を可能としているという、これまでの常識を覆す興味深い結果を得ました。今後は、Ca2+濃度の上昇に関与するチャネルの同定や、Ca2+による鞭毛波形の制御機構を明らかにすることで、長年の謎であった精子運動の制御システムが解明されることが期待されます。
また、精子走化性には種特異性があり、ホヤにおいても精子誘引物質SAAFの構造は種によって微妙に異なることがわかっており、さらにウニの精子走化性のメカニズムとは大きく異なることが明らかとなりました。今後は、受精の分子機構がどのように進化し種分化してきたかを解明するため、SAAFとPMCAの相互作用に種特異性があるかどうかを調べ、さらには他の動物とのシグナル系の比較を進めて行く予定です。
この記事を書いた人

-
吉田 学(写真左)
東京大学大学院理学系研究科附属臨海実験所 准教授
東京大学大学院理学系研究科動物学専攻博士課程修了 博士(理学)
日本学術振興会特別研究員、理化学研究所基礎科学特別研究員、東京大学大学院理学系研究科 助手、JST ICORPカルシウム振動プロジェクト研究員、等を経て2009年7月より現職。
研究分野は生殖生物学・細胞生物学。海産無脊椎動物から哺乳類まで幅広く受精の研究を行い、受精の分子機構が種分化する過程を解明したいと考えています。
吉田 薫(写真右)
桐蔭横浜大学医用工学部生命医工学科 准教授
東京大学大学院理学系研究科生物科学専攻博士課程修了 博士(理学)
聖マリアンナ医科大学博士研究員、桐蔭横浜大学先端医用工学センター助教、等を経て2016年4月より現職。
研究分野は生殖生物学・生化学。ヒトを中心に細胞外因子による精子の運動・受精能獲得調節機構の解明に取り組んでいます。
この投稿者の最近の記事
 研究成果2018年12月26日精子はどのように卵を見つけるか? – ホヤが受精するときの繊細な認識機構
研究成果2018年12月26日精子はどのように卵を見つけるか? – ホヤが受精するときの繊細な認識機構


