カブトムシの角の3D形態形成 -「折り畳んだ状態」で作り、脱皮と共に「展開する」
昆虫の「折り畳み」と「展開」による形態形成
蛹からチョウへ、ヤゴからトンボへ、芋虫からカブトムシへ、と昆虫は脱皮を介して短時間で劇的にその姿を変化させます。これは我々哺乳類が徐々に成長していくのと対照的です。なぜ昆虫は一瞬のうちにその姿を変化させることができるのでしょうか?
昆虫の体の表面は、硬いクチクラと呼ばれる外骨格に覆われています。昆虫はこの硬い外骨格のおかげで、捕食や病原菌感染、乾燥・加湿などの化学的変化への抵抗性を持っています。しかしその反面、硬く、伸張性に乏しい外骨格があることで体の成長は制限されています。そのため、昆虫は脱皮により、その都度古い外骨格を脱ぎ捨てることで成長します。
脱皮に先立って、新しい外骨格が古い外骨格の内側に形成されます。ここでポイントとなるのが、新しい外骨格は「皺々に折り畳まれた状態」で形成されることです。脱皮の際に、新しい外骨格の折り畳みが展開されることで、脱皮前よりも大きく成長することができます。もしも新しい外骨格に「折り畳み構造」がなければ、マトリョーショカ人形のごとく、脱皮のたびにどんどん小さくなってしまうことでしょう。
新しい外骨格を「折り畳んだ状態で作り、脱皮の際に展開する」というのは、昆虫で普遍的に見られる成長様式といえます。しかし、これまで「折り畳んで作る」という現象についてはあまり注目されていませんでした。これは、古い外骨格の内側という狭い空間で(元のものより大きな)新しい外骨格を作るのだから、受動的に自然と皺々の折り畳み構造ができてしまうと考えられてきたためと思われます。ところが、どうやら昆虫は能動的な発生プログラムとして、特定の折り畳み構造を作っているようなのです。
カブトムシの「角」は皺々に折り畳まれて作られる?
幼虫期のあいだは1回の脱皮ごとの形態変化が小さく、新しい外骨格の折り畳み度合いは比較的単純です。しかし、幼虫から蛹への脱皮の際には、一回の脱皮で大きく異なる姿へと変化するため、このとき新しく作られる構造では非常に複雑な外骨格の折り畳みが見られます。
カブトムシの突出した「角」はその典型的な例といえます。蛹で出現する角は、蛹化に先立ち幼虫の頭部内で幾重にも折りたたまれた「角原基」として形成され、蛹化時に体液が送り込まれることで展開され、細長く先端に分岐構造を持つ「角」となります(下図)。

その角原基やその形成過程を観察すると、皺の間隔に規則性があること、角の先端の分岐構造は馬蹄円状の皺によって規定されていること、折り畳み構造の形成順序は個体ごとにほぼ一定であることなどがわかりました。これらの現象は、皺々の折り畳み構造は、狭いスペースで角原基が発達するため「勝手にできてしまう」わけではなく、発生プログラムが能動的に作っていることを示唆しています。
展開後の最終的な立体構造など知る由もない角原基の細胞群集が、なんらかのルールに従って、分裂し移動し変形することで複雑な折り畳み構造を作り、最終的に「あとは展開するだけでOK」という状態になるというのは、単純に不思議なことではないでしょうか?
我々の研究グループは、カブトムシの角原基の折り畳み形成の謎を解くために、2015年ころから研究を開始しました。5年ほどかけて折り畳み構造を形作る発生メカニズムの一端を明らかにし、これまでに3本の論文を発表してきました。ここでは、これまでにわかったことをご紹介したいと思います。
角の3次元構造は、角原基の折り畳み構造で決まっているか?
まず、我々が取り組んだのは、「角原基の折り畳みを展開するだけで細長い角になる」という研究の前提の確認です。どういうことかといいますと、研究を始めた段階では、本当に折り畳みの展開だけで最終形への変化が起こるのかは確かめられておらず、展開に伴って細胞が増殖したり、原基がゴムのように伸張したりする可能性が残っていました。
そこで私たちは、生物を用いた実験およびコンピューターシミュレーションによる検証を行いました。その結果、細胞の増殖や移動、原基表面の局所的な伸長などが起こらない条件下でも、角原基の折り畳みを展開さえすれば最終形への変化が起こることが確認されました(下図)。

右)実際の原基を元に計算機上に構築した「バーチャル原基」の場合でも、正常な角形態に変化する。このシミュレーションでは、原基表面の伸張が起こらない条件で変化を起こしているため、純粋に折り畳みの展開だけでこの変化を説明できる。
(Matsuda et al. 2017 より改変して引用)
つまり、角原基の折り畳み構造が完成した段階で、展開後の角の形態は一義的に決まっていると明らかにできました。よって、折り畳み構造形成こそが、角の3次元形態形成であるといえます。
「折り畳み構造」を作る分子機構を検証
では、折り畳み構造の形成はどのような発生プログラムで起こっているのでしょうか? 先行研究により、カブトムシではすでにいくつもの角形成に関わる遺伝子が同定されています。これらの遺伝子の機能を阻害(ノックダウン)することで、角の形態をさまざまに変化させることができます。角の形態が変わるということは、角原基の折り畳み構造になんらかの変化が起こっているはずです。どのような遺伝子をノックダウンしたとき、角原基の折り畳み構造にどのような影響があるのかがわかれば、どのようなメカニズムが折り畳み構造形成に寄与しているのかわかるのではないか、と我々は考えました。
これまでにいくつかの遺伝子に注目して、角形態の変化と折り畳み構造の変化を解析した結果、どうやら一口に「折り畳み」といっても、そこにはいくつかの階層性が存在することが明らかになってきました。どういうことかというと、たとえば細胞極性の制御に関わる遺伝子である dachsous という遺伝子をノックダウンした個体では、角が太短くなることが知られています(下図A)。dachsous のノックダウン個体の角原基を見てみると、原基の形自体が大きく変わっている一方、表面の皺構造に大きな影響は見られませんでした(下図B、C)。
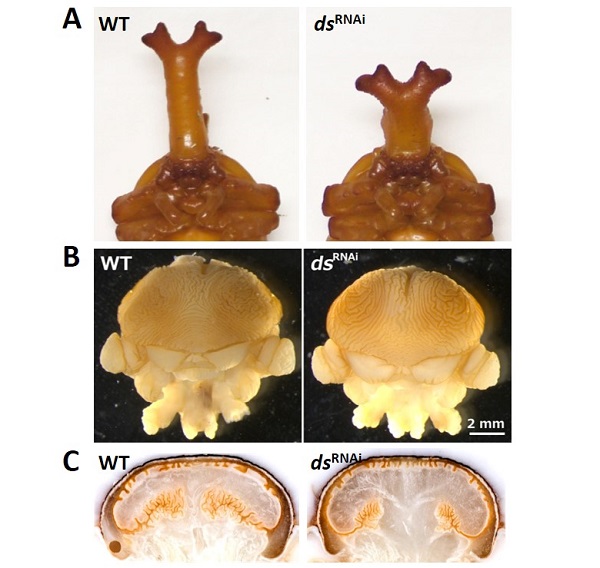
B)左:通常のオスの角原基、右:dachsous 遺伝子をノックダウンしたオスの角原基。dachsous 遺伝子をノックダウンしても、原基表面の皺構造に大きな変化は見られない。
C)左:通常のオスの角原基の断面、右:dachsous 遺伝子をノックダウンしたオスの角原基の断面。dachsous 遺伝子をノックダウンしてた個体では、原基全体の形(断面がキノコ状)が変化していることがわかる。
(Adachi et al. 2018 より改変して引用)
一方、細胞間シグナル伝達などに関わる遺伝子である Notch という遺伝子や、細胞分裂の制御に関わる遺伝子である Cyclin E という遺伝子をノックダウンしたときには、原基の形状に大きな影響は見られなかった反面、表面の皺に影響が見られました。
これらの結果から、角原基の折り畳みは「原基の全体の形状を決める大まかな折り畳み」と「原基表面の細かい皺状の折り畳み」に分けられ、(少なくとも部分的には)異なるメカニズムで作られていることがわかります。
さらに、表面の皺への影響は、Notch と Cyclin E のどちらの遺伝子をノックダウンするかで異なっており、Notch のノックダウンでは皺の深さが浅くなる一方、皺の方向は変わりませんでしたが、Cyclin E では逆に皺の深さは変わりませんでしたが、皺の方向に大きな乱れが見られました。つまり、表面の皺状の折り畳みについては、皺の深さを決めるメカニズムと皺の方向を(パターン)を決めるメカニズムは(少なくとも部分的には)独立しているといえます(下図)。
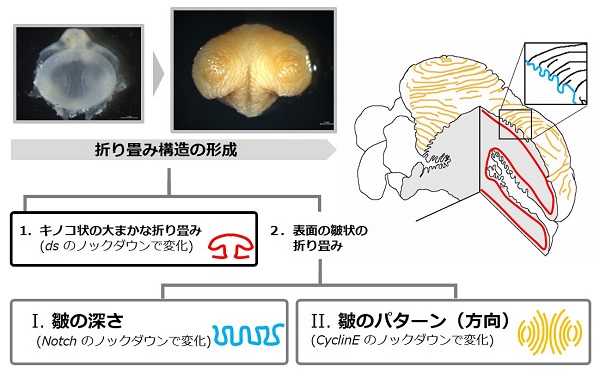
(Adachi et al. 2020 より改変して引用)
おもしろいのは、大まかな折り畳み、あるいは表面の皺の深さや方向のどれを変えても最終的な角の形態は変化するところです。現在、これらの遺伝子を手掛かりに、どのような細胞の挙動が変わることで、どう折り畳みの変化につながるのかや、他の昆虫種で見られる立体構造(クワガタムシの大顎や、ツノゼミのツノ、など)の折り畳みについても同じことが起こっているのかなどを明らかにすることを通して、昆虫で見られる「折り畳み」を介した形態形成の理解を目指しています。
参考文献
・Matsuda K*, Gotoh H*, Tajika Y, Sushida T, Aonuma H, Niimi T, Akiyama M, Inoue Y, Kondo S. Complex furrows in a 2D epithelial sheet code the 3D structure of a beetle horn. Scientific Reports, 7: 13939. 2017 https://doi.org/10.1038/s41598-017-14170-w
・Adachi H, Matsuda K, Niimi T, Inoue Y, kondo S, Gotoh H. Anisotropy of cell division and epithelial sheet bending via apical constriction shape the complex folding pattern of beetle horn primordia. Mechanisms of Development, 152: 32-37. 2018 https://doi.org/10.1016/j.mod.2018.06.003
・Adachi H, Matsuda K, Niimi T, Kondo S, Gotoh H. Genetical control of 2D pattern and depth of the primordial furrow that prefigures 3D shape of the rhinoceros beetle horn. Scientific Reports, 10: 18687. 2020. https://doi.org/10.1038/s41598-020-75709-y
この記事を書いた人

-
国立遺伝学研究所 特任研究員
博士(環境科学)。 1984年北海道生まれ。北海道大学大学院 環境科学院 生物圏科学専攻 博士後期課程修了後、海外学振特別研究員(於ワシントン州立大学)、名古屋大学大学院 生命農学研究科 特任助教などを経て現職。次のポジション求職中!(2022年3月で任期切れ)
この投稿者の最近の記事
 研究成果2020年12月9日カブトムシの角の3D形態形成 -「折り畳んだ状態」で作り、脱皮と共に「展開する」
研究成果2020年12月9日カブトムシの角の3D形態形成 -「折り畳んだ状態」で作り、脱皮と共に「展開する」 研究成果2017年2月28日クワガタムシの発達した大顎を形作る遺伝子とは?
研究成果2017年2月28日クワガタムシの発達した大顎を形作る遺伝子とは?


