植物は葉緑体に貯えた「たくさんのDNA」を自己分解してリン栄養を得る – 細胞内共生から生じた生存戦略
植物のDNAとリン栄養の関係
植物は葉緑体で行われる光合成によって大気中の二酸化炭素から有機物を作りますが、窒素(N)やリン(P)、カリウム(K)などの養分は、ヒトや動物のように外部から吸収して利用します。N、P、Kは植物の三大栄養素として肥料にも使われています。
リン(有機リン)は細胞内でリン酸として利用されており、リン酸を最も多く含む物質はDNAやRNAなどの核酸です。DNAは親からの遺伝情報を受け継ぐ生命の維持に必須の遺伝物質ですが、ヌクレオチドとしてリン酸を多く含むことから、DNAを分解して再利用すれば栄養素としても有効だと考えられます。たとえば、DNAの断片化は細胞死の指標とされますが、分解されることの生理的意義や、リン栄養との関連についてはよくわかっていませんでした。
最近筆者らは、植物の葉緑体に着目したところ、DNAが分解されてリンの再利用に役立っているという現象を明らかにしました。どうやら植物はDNAを、遺伝子としてだけでなく、「リンの貯蔵物質」として、リン欠乏などの外的環境に応答して生き残るための戦略に用いているようです。ここでは、葉緑体のDNAと、DNA分解によるリン貯蔵機能について紹介します。
細胞内共生に由来する葉緑体には、DNAが「たくさん」ある
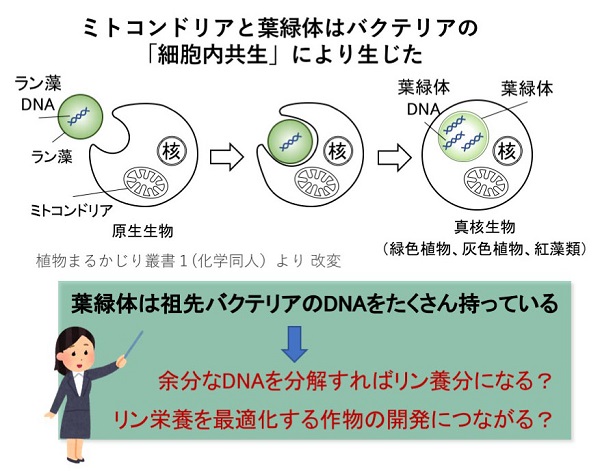
今から半世紀以上前、分子生物学黎明期である1960年代に、細胞内小器官(オルガネラ)であるミトコンドリアと葉緑体(プラスチド)にDNAがあることがわかりました。それまでにわかっていた核の染色体DNAと区別するため、ミトコンドリアやプラスチドのDNAは「オルガネラDNA」と呼ばれます。オルガネラDNAの存在は、葉緑体とミトコンドリアが、それらの祖先である光合成細菌(シアバクテリア)とαプロテオバクテリアの共生に由来して真核生物が進化したという、「細胞内共生説」を支持する強力な証拠になっています。
葉緑体DNAにはシアノバクテリアに由来する100個ほどの遺伝子があり、これらが転写・翻訳されることでタンパク質を合成します。しかし、葉緑体を構成するタンパク質は3000個以上あり、残りのタンパク質を作る遺伝子は、共生進化の過程ですべて葉緑体DNAから染色体DNAに移行してしまっています。細胞内共生から細胞の一部になった葉緑体は、進化の過程でほとんどのDNAを核のDNAに転移させ、自律性を失ったと考えられます。
では、なぜ全部の遺伝子を核に転移せず、一部の遺伝子だけを葉緑体に残す必要があったのでしょうか。可能性としては、光合成反応などの特に重要な遺伝子だけを残している、あるいは、すべてのDNAを転移する進化の途上にある、とも考えられます。さらに、光合成をしなくなった寄生植物のプラスチドにもDNAがあることから、葉緑体自体がDNAをもつ必要性があることも考えられます。
また、葉の細胞は葉緑体DNAを非常にたくさん持っています。通常の体細胞では核DNAは2コピーであるのに対し、葉緑体DNAはときに1000コピー以上になることもわかっています。葉緑体は遺伝子数が圧倒的に少ないのとは対照的に、DNAのコピーの量は多く、全DNA量の30%以上を占めることもあります。なぜ、葉緑体はわざわざこれほどの量の葉緑体DNAを持っているのでしょうか。
オルガネラDNAを分解する酵素「DPD1ヌクレアーゼ」
葉緑体にあるDNAの量が変動するかどうかについては、普遍的に決まった量が存在する、あるいは分解されてなくなる、という食い違った意見をアメリカとドイツの研究者らが提唱し、論争が10年以上続いていました。この論争を決着するデータとして筆者らは、「DPD1ヌクレアーゼ」というDNA分解酵素を発見し、DNAが分解されていることを見つけました。
この研究では、シロイヌナズナというモデル植物で、花粉ができるときにオルガネラDNAが分解されると仮定し、DNAが分解されない突然変異体を見つけて詳しく調べることにしました。その結果、DPD1ヌクレアーゼをみつけることができました。
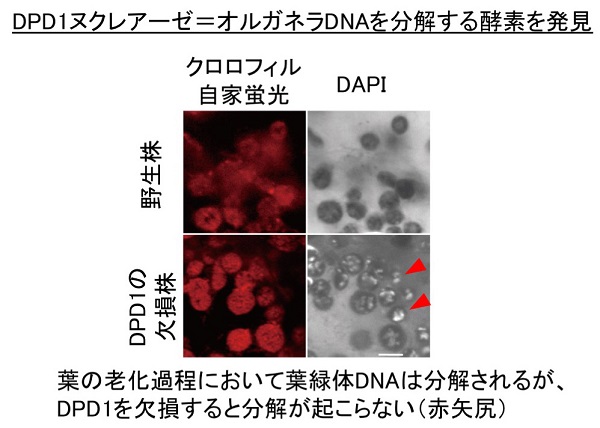
DPD1ヌクレアーゼが発現する場所を詳しく調べてみると、花粉だけではなく、葉が老化するときにも著しく発現が誘導されることがわかりました。ではなぜ、DNAが分解されるのでしょうか。その疑問を解決するために、「葉の老化」とリンとの関わりについて掘り下げてみました。
植物は葉緑体DNAを自己分解してリンをリサイクルしている
動物のように動いて自分の栄養を探して摂取することをあきらめた植物は、地に根を張って養分を吸収し、光合成で成長しながら陸上で進化し繁栄してきました。そのため、不足する養分に適応するためのさまざまな適応機能を発達させています。
「葉の老化」はそのようなプロセスのひとつで、植物は自分が光合成などに使った高分子化合物を分解して転流させ、「再利用」しています。たとえば、実りの秋の田んぼが一斉に黄金色になり、稲穂がこうべを垂れるのは、葉の老化で養分が効率よく稲穂に転流して使われるからなのです。
筆者らは、DNAが壊れるのもこの再利用のひとつの過程であると考えました。特に、DNAがリンを多く含むことを考えると、DPD1ヌクレアーゼによる葉緑体DNAの分解が、リン栄養の供給に関係すると予想しました。結果は予想どおりで、植物をリン欠乏させた状態で栽培すると、DNA分解が起こらないシロイヌナズナ変異体は典型的なリン欠乏症状を発症し、リンのリサイクルが停滞して種子を作る能力も低下してしまいました。
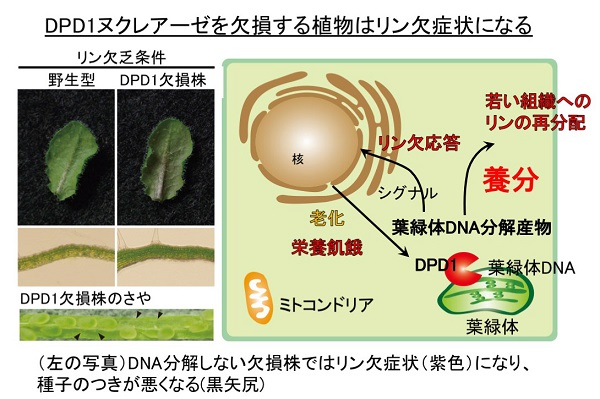
植物をリン欠乏の状態にすると、植物は外部のリンを取り込んだり、リン脂質を再構成する応答反応が起こることが知られていますが、DNA分解が起こらないと、この応答反応も低下することがわかりました。つまり、共生進化の痕跡として葉緑体にたくさん残しているDNAは、リン貯蔵の役割もしていて、リン欠乏などの状況に応じてこのDNAを再利用して、その分解作用にDPD1ヌクレアーゼがはたらいているということがわかりました。
この現象は種子植物に普遍的であり、落葉樹(ポプラ)が葉の落葉でリンを再利用するときにも、DPD1ヌクレアーゼによって葉緑体DNAが積極的に分解されていることもわかりました。
葉緑体DNAはリン貯蔵物質モデルにあてはまる
ワトソンとクリックが1953年にDNAの二重らせん構造を解明し、DNAが遺伝情報物質とわかり、現在ではDNAの研究があらゆる分野で進んでいます。DNAは、実はこの発見に遡る1860年代にドイツ・チュービンゲン大学の化学者フリードリヒ・ミーシャーにより初めて単離されています。ミーシャーは、患者の膿の細胞から単離した物質(DNA)を当時「ヌクレイン」と名付けました。
ミーシャーは、この「ヌクレイン」がタンパク質とは異なり多量のリンを含む物質であることを見つけ、リンの細胞内貯蔵に関わる可能性を述べています。しかし、これまで遺伝子の研究が進む一方で、リン貯蔵についてはあまり調べられてきませんでした。今回の研究は、細胞内共生により維持されている植物のDNAについて、このようなリン貯蔵の機能があることを明らかにしています。また、細胞内共生による進化とDNAの疑問にもひとつの答えを提示しています。植物はDNAの一部を葉緑体に残すことで、葉緑体に大きな核酸のプールを形成し、リン貯蔵の役割を担わせることで、リン栄養の枯渇に備えているのかもしれません。
現在、リン肥料は主に天然のリン鉱石から作られていますが、21世紀になりリン鉱石の埋蔵量が懸念され、今世紀中に枯渇するのではとも危惧されています。今回の研究により、葉緑体におけるDNAとリン再利用の関係が明らかとなったので、オルガネラDNA量をコントロールすることでリン利用効率の向上した作物の育成にもつながることが期待されます。

参考文献
この記事を書いた人

-
坂本 亘(写真左)
岡山大学資源植物科学研究所教授。1990年東京大学大学院農学系研究科修了(農学博士)。コーネル大学ボイストンプソン植物研究所ポスドク後、1993年より現在の研究所に在籍。専門は植物生理学、遺伝学。所属研究所では、光環境適応研究グループのリーダーとして、光合成と葉緑体分化の環境適応をさまざまな角度から研究しています。大学の細胞学で細胞内共生に魅せられてから、オルガネラの遺伝学と生理学を志し、気がついたらいい年になりましたが、これからも光合成と葉緑体の不思議を追い続けたいと思います。
高見 常明(写真右)
岡山大学資源植物科学研究所技術専門職員。2010年九州大学大学院生物資源環境科学府修了(博士(農))。九州大学理学府でのポスドクを経て、2012年6月より現在の研究所で光環境抵抗研究グループに技術職員として所属。専門は光合成と植物生理学。こつこつと植物の持つ能力を研究していけたらと思っています。