視覚進化の新モデル – 哺乳類が失った”ピノプシン”からたどる脊椎動物の暗所視と色覚
眼と視覚の進化
動物にとって、眼から得られる視覚の情報は外界の変化をとらえるうえで非常に重要です。特にヒトでは、周りから得られる感覚情報の多くが視覚を通じて得られるといわれ、脳の約半分が視覚の情報処理に関わっています。ヒトなどの脊椎動物はカメラ眼と呼ばれる眼をもち、ハエなど昆虫類が持つ複眼とは見た目が大きく異なっています。
かつて、チャールズ・ダーウィンもさまざまな形態をもつ眼の進化に大いに興味を持っていたことが、その著書『種の起源』の記述からもうかがえます。そして最近では、眼の形態的な違いだけでなく、眼の中で機能する分子(タンパク質)に着目した研究も進んできました。
ヒトなどの脊椎動物は高度に発達した視覚を持ち、真夏の太陽がさんさんと照りつける明るい場所でも、月の出ていない星空の下のような暗い場所でも、ものの形を認識できます(明所視と暗所視)。また、明るい場所では色も認識できます(色覚)。この視覚機能を支えているのが、ロドプシンに代表される光センサー(光受容タンパク質)です。
眼の奥、網膜にある視細胞の中でこの光受容タンパク質が光を受け取ると、光シグナルは視細胞の電気応答に変換されます。この電気応答が最終的に脳へと伝達され、”見えた”と実感できます。約5億年前、カンブリア爆発と呼ばれる生物の多様化の時期までに他の動物群と分岐した脊椎動物の祖先は、明暗を感じる程度の眼点しか持っていなかったと考えられます。では、脊椎動物の祖先はどのようにして明所視・色覚と暗所視といった高度な視覚を手に入れたのでしょうか。
脊椎動物の明所視・色覚と暗所視
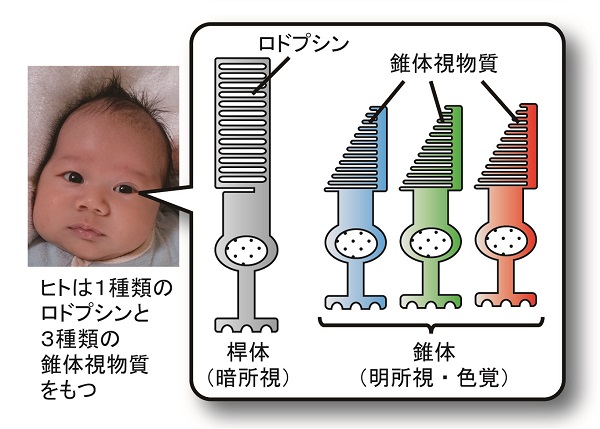
多くの脊椎動物の眼には、桿体と錐体と呼ばれる2種類の視細胞があります。桿体は薄暗がりで働き(暗所視)、錐体は明るいところで働きます(明所視)。2種類の視細胞では機能する光受容タンパク質も異なっており、桿体ではロドプシン、錐体では錐体視物質が働いています。ヒトを含む多くの脊椎動物は、吸収する光の波長が異なるいくつかの錐体視物質(ヒトでは、赤色光・緑色光・青色光それぞれを受容する3種類)をもつ一方、ロドプシンは1種類しか持ちません。そのため、昼間(明所視)は色を見分けられますが、夜(暗所視)になると色がわからなくなります。
このような明所視・色覚と暗所視がどのように獲得されたのかという疑問は、「周りのものがどう見えるのか」というヒトにとって非常に重要な問題につながるため、大きな議論の的となってきました。明暗視に比べて色覚の方が複雑な情報処理を必要とすると考えられるため、1990年代初頭までは、明るいところで働く視覚(明所視)と暗いところで働く視覚(暗所視)が分岐した後に、明所視から色覚を獲得したと推測されていました。
しかし約25年前、ロドプシンや錐体視物質のアミノ酸配列を調べることで、実験的に進化の道筋が明らかにされました。ロドプシンや錐体視物質のアミノ酸配列が明らかになると、その比較からどちらが先に進化したのか(先祖型なのか)を推定することができます。
その結果、脊椎動物の祖先は吸収する光の波長の異なる複数の錐体視物質を先に創り出し、その後にロドプシンを創り出したことがわかりました。これは、先に明所視から色覚の進化があり、その後に暗所視の進化が続いたということを示し、それまでの推測とは逆の結果です。この進化モデルは当時非常にインパクトがあり、今では多くの教科書でも記述されるほど広く知られるものとなっています。今回私たちは、この定説を修正する新たな発見をしました。
眼で機能するピノプシン
私たちは、ロドプシンや錐体視物質に近いアミノ酸配列を持つ光受容タンパク質、”ピノプシン”に注目しました。ピノプシンはニワトリの脳内から見つかった光受容タンパク質で、ロドプシンの類似タンパク質が眼だけでなく脳内でも働いていることが実証されたため注目されていました。その後、ピノプシンは他の鳥類や爬虫類でも脳内で機能し、眼では機能していないことが示されたため、もっぱら視覚以外の光受容機能(たとえば、光環境の変化から時刻を知る)に関わると考えられていました。
そこで今回私たちはまず、鳥類や爬虫類だけでなく、脊椎動物の進化の初期に系統的に分かれたと考えられる軟骨魚や古代魚について調べ、これらのゲノムにもピノプシン遺伝子が存在することを見いだしました。そして実際に軟骨魚(サメ)や古代魚(ガー、チョウザメ、ポリプテルス、ハイギョ)を使って調べると、ピノプシンは脳内だけでなく、眼にも存在することがわかりました。また、両生類のカエルでもピノプシンは眼にも存在していました。さらに詳しく調べると、ピノプシンはロドプシンと同様に桿体で機能し、そのタンパク質の性質もロドプシンと非常によく似て暗所視に向いていることがわかりました。
これらの結果から、ピノプシンは脳内で働くだけでなく、ロドプシンと同様に薄暗がりで働く桿体用の光受容タンパク質として、脊椎動物の祖先により獲得されたと考えられます。そしてその後、脊椎動物の進化に伴って脳内での光受容機能に特化するように変化したと想像できます。
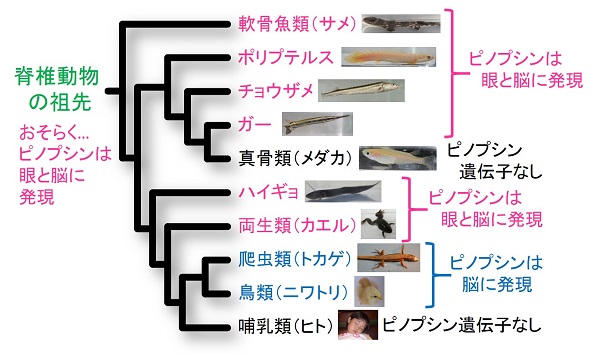
脊椎動物の祖先はピノプシンを眼と脳で使っていたと考えられる。そして、爬虫類・鳥類への進化の過程で脳でのみ使うようになり、哺乳類ではピノプシンを使って脳で光受容することをやめたため、ピノプシン遺伝子を失ったと考えられる。
ピノプシンのアミノ酸配列をロドプシンや錐体視物質と比べると、複数の錐体視物質が創り出される最初期にピノプシンも創り出されていることがわかります。つまり、この時点で脊椎動物の祖先はすでにピノプシンを使って暗所視を実現できていた可能性が高く、その後ロドプシンを創り出すことでより高度な暗所視を獲得したと推定できます。これらのことから、「色覚の獲得が先で、暗所視の獲得が後」という定説を修正し、「色覚の獲得と並行して暗所視の獲得が2度起こった」と推測できます。
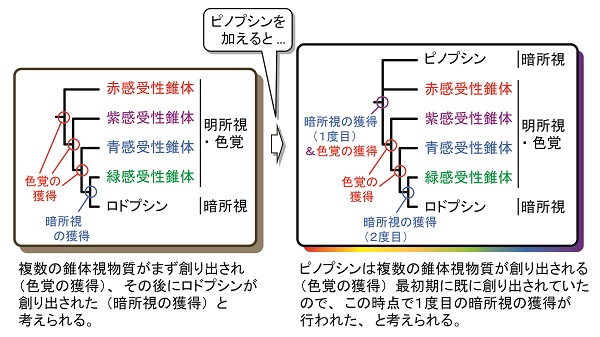
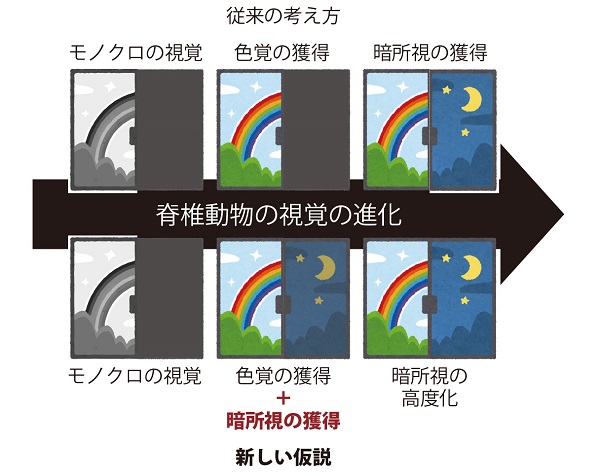
従来のモデルでは、脊椎動物の先祖において明所視から色覚が先に獲得され、暗所視の獲得が続いたと考えられている。しかし新モデルでは、色覚の獲得と並行して暗所視の獲得が2度起こったと考える。
視覚進化の謎は続く
今回解析したピノプシンは現存の魚類や両生類では眼と脳で働きますが、爬虫類や鳥類では脳だけで働いています。そして、哺乳類ではピノプシンを使って脳で光受容することをやめてしまったため、ピノプシン遺伝子自身をなくしています。また、魚類や爬虫類、鳥類は4つの錐体視物質を使って4色性色覚を持つものが多いものの、哺乳類は霊長類を除くほとんどが2つの錐体視物質を使う2色性色覚です。つまり、哺乳類は進化の過程で錐体視物質の遺伝子もなくしています。
哺乳類がこのように光受容タンパク質のいくつかを失っているのは、その祖先が夜行性であったためと推測されています。逆にいうと、哺乳類以外の脊椎動物はヒトよりも多種類の光受容タンパク質を持つため、より多様な形で光を利用して周りから情報を得ている可能性があります。たとえば、多くが夜行性のカエルは夜でも色を認識できるといわれています。最近私たちは、カエルがもつこの「暗所での色覚」という特殊能力をもたらす分子的なメカニズムを明らかにすることに成功しています。しかし、まだまだ視覚進化には謎がいっぱいです。動物たちが、どのようにして視覚機能を独自に進化させ多様な環境に適応しているのか、興味が尽きません。
参考文献
この記事を書いた人

-
山下高廣(写真左)
京都大学大学院理学研究科助教。2002年、京都大学大学院理学研究科博士後期課程修了 博士(理学)。日本学術振興会特別研究員(PD)を経て、2004年より現職。ロドプシンなどのGタンパク質共役型受容体に始まる細胞内情報伝達系の多様な機能とその分子メカニズムを研究している。
佐藤恵太(写真中)
岡山大学大学院医歯薬学総合研究科助教。2012年、京都大学大学院理学研究科博士後期課程修了 博士(理学)。同大学大学院理学研究科研究員を経て、2016年より現職。生物が光をどのように利用するか、光が生物にどのような影響を与えるかに興味をもち、特に動物の非視覚性光受容の分子メカニズムについて研究している。
七田芳則(写真右)
立命館大学総合科学技術研究機構客員教授。1979年、京都大学大学院理学研究科博士後期課程修了 博士(理学)。同大学大学院理学研究科助手、助教授を経て1998年同大学大学院理学研究科教授。2017年より現職(京都大学名誉教授)。



