テントウムシの斑紋パターンはどう決まる? – ひとつの遺伝子が多様な斑紋をつくる仕組み
ナミテントウの斑紋はメンデルの法則に従う
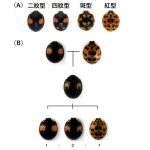
テントウムシは世界で6,000種、日本で180種が同定されており、種によって決まった斑紋をもちます。なかには斑紋多型とよばれる同じ種であっても遺伝的に決まった複数の斑紋をもつものがあります。ナミテントウの斑紋多型は特に顕著で、200以上もの斑紋が知られています。

このような多様な斑紋が生じる遺伝様式を明らかにするため、我が国の研究者を中心に遺伝学的な交配実験が古くから行われてきました。その結果、斑紋型についてホモ接合体の親を交配した場合、その子の斑紋は必ず両親の斑紋を重ね合わせた斑紋になることが明らかになりました。両親の斑紋の重ね合わせで、赤い領域と黒い領域が重なる領域は、必ず黒色になります。次に、その子同士を交配した孫の世代の斑紋は、メンデルの遺伝の法則に従い両親の斑紋と子の斑紋が必ず1:2:1になるように現れます。
この結果は、どの斑紋の交配においても当てはまります。したがって、ナミテントウの多様な斑紋は、ゲノム中のたったひとつの遺伝子のなんらかの違いによってもたらされることが予想されました。しかしながら、ナミテントウはもちろんのことその他のテントウムシにおいても、斑紋を作るために必要な遺伝子はまったく不明でした。そこで、私たちの研究室では、最新の手法を駆使してナミテントウの斑紋を作る遺伝子を明らかにするプロジェクトに取り組んできました。
ナミテントウの斑紋の色素はどのように分布している?
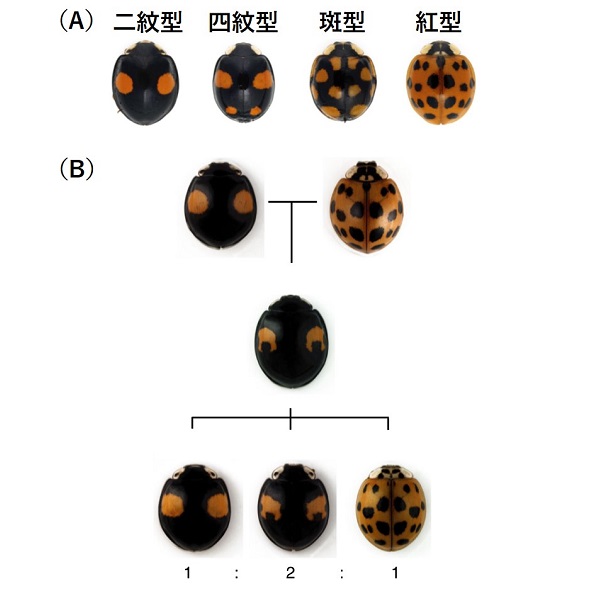
まずはじめに、翅の斑紋が発生過程においてどのように生じるのかを調べました。ナミテントウの翅は、蛹から脱皮した羽化直後は全体が黄色で、黒いところはまったくありません。羽化して2時間後に、黒くなる部分はうっすらと黒く色付き始め、その後数時間で黒色になります。
興味深いことに、羽化直後の黒くなる以前の翅をよく観察すると、翅は全体が均一な黄色ではなく、将来赤くなる部分は黄色ですが、黒くなる部分には黄色い色素がなく、白っぽく見えました。つまり、両親の斑紋を重ね合わせた場合に赤い領域と黒い領域が重なるところでは黒色になりますが、これは赤い色素(カロテノイド)が黒い色素(メラニン)によって見えなくなっているのではなく、赤い領域と黒い領域は互いに重なることがないように、領域分けされていることを示しています。また、メラニンはテントウムシ自身が合成できますが、カロテノイドは合成することができず餌から取り込んだ色素が利用されます。
ここまでは翅という平面上での赤色色素と黒色色素の分布について見てきましたが、次に翅の断面における色素の分布について調べました。昆虫の翅は1枚のシート状の構造になっていますが、断面を見ると背側と腹側のそれぞれ一層の細胞から作られることが知られています。そこで、翅の切片を作製して、翅の断面において、斑紋の色素がどのように分布しているのかを調べました。その結果、興味深いことに、黒い色素は背側の細胞で作られ、赤い色素は腹側の細胞に蓄積することが明らかになりました。つまり、背と腹の細胞間には何らかの相互作用があり、その結果背と腹で異なる色素が着色する領域が厳密に制御されて、斑紋のパターンが作られることが示唆されました。
ナミテントウの斑紋パターンを決定する遺伝子
次に、ナミテントウの斑紋形成に関与する遺伝子を見つけるため、翅形成の研究が最も進んでいるショウジョウバエの知見を参考に、ナミテントウの斑紋形成に関わっている可能性がある遺伝子を選びました。ナミテントウからこれらの遺伝子を単離して、遺伝子の機能を阻害する実験を行いました。この実験では、遺伝子の機能がなくなることにより、斑紋のパターンが変化する遺伝子をしらみつぶしに調べていきました。
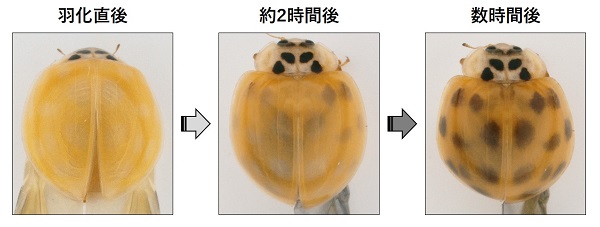
幸運にも、最初に調べた10個の遺伝子のなかに、斑紋のパターンが変化する遺伝子を見つけることに成功しました。この遺伝子は、パニア(Pannier)とよばれる転写因子をコードする遺伝子であり、さまざまな遺伝子の発現を制御する機能をもっています。パニア遺伝子の働きをなくすと、翅の斑紋の黒い領域が赤い領域に変化して、翅全体が赤くなりました。
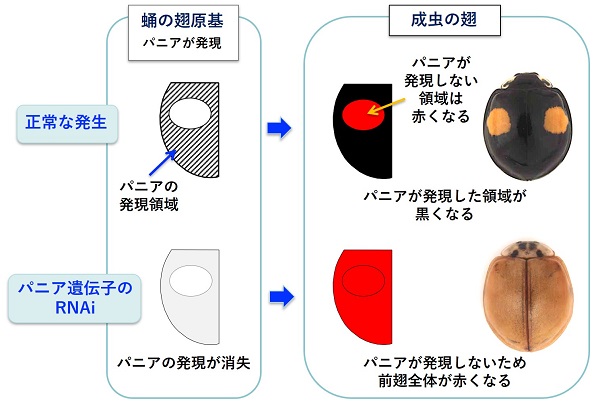
さらに、パニア遺伝子は、蛹において翅に色素が着色する直前のステージで、将来黒くなる背側の細胞で働き始めることが判明しました。これらの結果は、前述した色素の着色特性によく一致しています。つまり、パニア遺伝子が働く背側の細胞では黒い色素であるメラニン形成が生じ、一方その腹側にある細胞では赤い色素であるカロテノイドの着色が阻害されると考えられます。従って、パニア遺伝子の働きがなくなると、メラニンが形成されず、同時にカロテノイドの着色が生じることになります。以上の結果から、パニア遺伝子は、たったひとつの遺伝子の働きで翅全体のパターンを決定する興味深い機能をもつことが明らかになりました。
ナミテントウの斑紋多型をもたらす原因遺伝子を特定
次の解決すべき問題は、果たしてパニア遺伝子は斑紋多型をもたらす遺伝子であるのかということです。このことを明らかにする際に、ナミテントウのゲノム情報の利用が不可欠でした。ナミテントウの二紋型のゲノム解読を行ったところ、良好な解読結果が得られ、ゲノムサイズは423Mbと判明しました。このゲノム情報を利用して、ゲノム全体にわたる連関解析を行うことにより、斑紋多型の原因遺伝子が存在する領域をゲノム中の0.66~2.1Mbまで絞り込むことができました。
この領域には、19個の遺伝子が存在していましたが、遺伝子の発現解析から斑紋の形成過程の黒色領域で発現が有意に高くなる遺伝子は唯一パニア遺伝子のみでした。したがって、パニア遺伝子は斑紋多型の原因遺伝子であることが明らかになりました。次に、斑紋型間でのパニア遺伝子の塩基配列の違いを特定するため、二紋型に加えて斑(まだら)型と紅型についてゲノム解読を行い、配列を比較しました。その結果、パニア遺伝子の配列のなかでタンパク質を作る配列には、斑紋型の違いを説明できる塩基配列の相違は存在せず、興味深いことに、斑紋型間の配列の違いは第1イントロンにあることが判明しました。
詳細にナミテントウのパニア遺伝子の第1イントロンを調べてみると、他の昆虫にはない特徴がいくつかありました。ひとつ目は、第1イントロンの配列が非常に長くなっていたことでした。2つ目は、第1イントロンの配列は斑紋型間で非常に多様性に富んでいました。3つ目は、この第1イントロンの配列には、染色体の逆位が複数回生じていました。一般に染色体に逆位が生じるとその領域の組換えが抑制されるため、異なる斑紋をもたらす配列上の相違が維持されていると考えられました。
パニア遺伝子の機能は他の昆虫でも保存されるか
少なくともショウジョウバエを含む他の昆虫ではパニア遺伝子が着色を制御する機能は報告されていません。それでは、着色機能の起源はどこまで遡れるのでしょうか。その謎に迫るため、手始めにナミテントウと同じテントウムシ亜科に属し、ナミテントウに比較的近縁なナナホシテントウについてパニアの機能を調べてみることにしました。その結果、ナナホシテントウのパニア遺伝子もナミテントウと同様に斑紋パターンを決定する機能をもっていることが明らかになりました。ナミテントウとナナホシテントウの共通祖先は、約3,390万年以上前に分岐したと考えられており、これら2種の共通祖先のパニア遺伝子は、すでに斑紋パターンを決定する機能をもっていたと考えられます。
今後は、ナミテントウのパニア遺伝子の第1イントロンのどのような配列の違いによって異なる斑紋パターンが作られるのかを明らかにしたいと考えています。また、翅に斑紋をもつ昆虫は多くいますが、それらの昆虫のパニア遺伝子にも今回見出された斑紋パターンを決定する機能があるのかを調べることにより、パニア遺伝子によって斑紋パターンがもたらされた昆虫の起源に迫りたいと思います。
参考文献
・Ando, T., Matsuda, T., Goto, K., Hara, K., Ito, A., Hirata, J., Yatomi, J., Kajitani, R., Okuno, M., Yamaguchi, K., Kobayashi, M., Takano, T., Minakuchi, Y., Seki, M., Suzuki, Y., Yano, K., Itoh, T., Shigenobu, S., Toyoda, A., Niimi, T. (2018) Repeated inversions within a pannier intron drive diversification of intraspecific colour patterns of ladybird beetles. Nat. Commun., 9, 3843.
・Komai, T. (1956) Genetics of Ladybeetles. Adv. Genet. 8, 155-188.
・Teruyuki Niimi, (2018) Nature Research Ecology & Evolution Community: Behind the Paper “Wing colour prepattern gene in ladybird beetles”
この記事を書いた人
-
新美輝幸
基礎生物学研究所 進化発生研究部門/総合研究大学院大学 教授。博士(農学)。1965年愛知県半田市生まれ。名古屋大学大学院農学研究科博士課程(後期課程)修了後、日本学術振興会特別研究員PD(名古屋大学)、日本学術振興会海外特別研究員(バーゼル大学)、名古屋大学大学院生命農学研究科助教、科学技術振興機構さきがけ研究研究者(兼任)を経て現職。昆虫の翅、テントウムシの翅の模様、カブトムシの角など昆虫が進化の過程で独自に獲得した新奇形質に着目して、昆虫の多様な形質をもたらす分子基盤および進化メカニズムを解明することを目指している。
安藤俊哉
自然科学研究機構・基礎生物学研究所・進化発生研究部門/総合研究大学院大学・基礎生物学専攻 助教。博士(生命科学)。1983年福島県生まれ、神奈川県育ち。東京大学理学部生物学科動物学コース卒業。同大学大学院新領域創成科学研究科先端生命科学専攻博士課程修了後、同専攻・特任研究員、理化学研究所CDB・研究員を経て2016年より現職。専門分野:発生遺伝学、進化生物学。カイコ・ショウジョウバエ・テントウムシ・蝶などのさまざまな昆虫が示す昆虫特有の特徴的な体づくりに関わる遺伝的基盤の分子レベルでの解明を目指している。