脳の回路はどう作られる? – 赤ちゃんマウスの神経回路発達プロセスを直接観察
赤ちゃんは日々驚くべきスピードで成長し、毎日違った表情を私たちに見せてくれます。ついこの前までベッドの上で寝転んでいただけだったのに、ふと気づいたら、這って、立って、歩いて、話して……。こうした行動は脳によってコントロールされています。脳の機能は神経細胞のネットワーク(神経回路)により担われており、神経回路は胎生期に遺伝的プログラムにより大まかに形成されたのち、主に出生後に使われながら再編され、成熟した回路になっていきます。赤ちゃんの驚異的な成長スピードはこの回路再編機構のなせるわざであり、また回路再編が適切に進まないことが自閉症などの発達障害の一因となることも示唆されています。しかし、新生児期に神経細胞が実際にどのように姿を変えて機能的な回路を形作っていくのかはほとんどわかっていませんでした。今回、私たちは生きた新生仔マウスの脳内で、特異的な回路を作るために神経細胞が形を変えていく様子を直接観察することに成功しました。
マウス脳の「バレル」構造
神経細胞は、(多くの場合)複数本存在する太くて短い樹状突起から情報を受け取り、1本の細くて長い軸索によって次の神経細胞の樹状突起へと情報を伝達します。したがって、樹状突起の形は神経回路の機能の根幹であり、発達期に神経細胞が適切な軸索に向けて樹状突起を展開する仕組みを知ることは、機能的な脳の神経回路が構築される機構の理解に不可欠です。この樹状突起展開の仕組みの解明のため、私たちはマウス大脳皮質体性感覚野を観察対象としました。
夜行性のマウスにとってヒゲの感覚は重要であり、ヒゲからの体性感覚(触覚)情報を処理する領域は大脳皮質のなかでもひときわ大きな割合を占めています。ヒゲ領域では1本1本のヒゲに対応する部分が組織学的に分離して、樽(barrel)のように見えることから「バレル」と呼ばれる構造を構成しています。
ヒゲからの情報は脳幹、視床を経由し、視床皮質軸索を介して大脳皮質のバレルへと届けられます。それぞれのバレルの内側(樽の中身にあたる部分)には1本のヒゲからの情報を伝える視床皮質軸索の末端がクラスターを形成しています。そしてバレル神経細胞の多くはバレルの縁に存在し、バレルの内側に向けて選択的に樹状突起を展開することで、1本のヒゲの情報のみを受け取っています。この樹状突起の方向性は生後1週間のうちに神経入力の影響下に形成されることが知られており、私たちはその仕組みに長期生体イメージングというアプローチで迫りました。
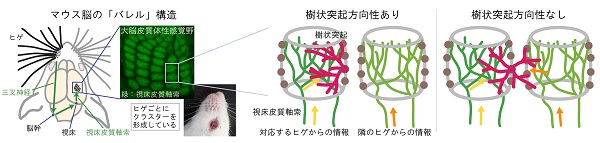
(中)樹状突起(赤)に方向性があることで、1個の神経細胞には1本のヒゲの情報のみが入る
(右)方向性が失われると回路が混線してしまう
生体イメージングへ向けた技術開発
生体イメージングのためには大脳皮質の神経回路を蛍光タンパク質で標識する必要があります。そのため、これまでに私たちは2つの技術開発を行いました。ひとつは、視床皮質軸索(TCA)で緑色蛍光タンパク質(GFP)を発現するTCA-GFPトランスジェニックマウスの開発です。視床皮質軸索を緑色に光らせることでバレルの位置を可視化できました。
もうひとつは、狙った脳領域の神経細胞を高輝度かつまばらに蛍光標識する方法(Supernovaシステム)の開発です。特殊な仕掛けのある2種類のベクターを胎仔期の脳に電気的に導入するだけの簡便な手法であり、これにより新生仔大脳皮質に高密度に存在する神経細胞のなかからごく少数だけを光らせることに成功し、樹状突起形態の詳細な解析が可能となりました。
そして、これらの技術と2光子励起顕微鏡(不透明な組織の深部を観察できる顕微鏡)の組み合わせにより、生後5日齢マウスのバレルの18時間にわたる生体イメージングに成功し、2014年に報告しました。これは哺乳類の脳内で樹状突起が動く様子を直接観察した世界で初めての例でした。今回はその観察技術をさらに発展させ、より幼い時期(生後3日齢)から、3日間に渡る長期生体イメージングに成功しました。その結果、神経細胞の樹状突起の方向性獲得・増強という大きな時間スケールで起こるイベントの解析が初めて可能となりました。
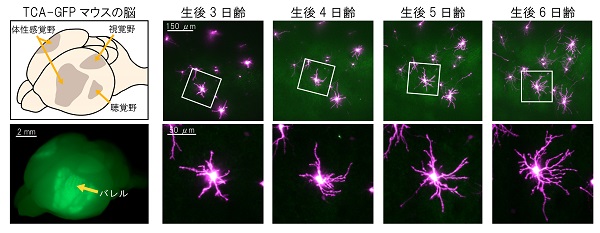
緑:TCA-GFPトランスジェニックマウスで可視化されたバレル
赤:Supernovaシステムによりまばらに標識されたバレルの神経細胞
※実際の撮影は8時間ごとに行われた
長期生体イメージングで明らかとなった樹状突起の成長のしくみ
長期生体イメージングにより同じ神経細胞の形態を繰り返し撮影した結果、新生仔期の脳でバレルの縁に位置する神経細胞ではバレルの内側でも外側でも、細胞体から短い樹状突起が新しく作られてはすぐに消えるというターンオーバーが繰り返されていることを発見しました。しかし、新しく作られた樹状突起のすべてが消えるわけではありません。一部は安定化し、「勝者」として複雑に成長していきました。重要なことに、こうした勝者は主に(適切な軸索の存在する)バレル内側に生まれた樹状突起のなかから現れていました。
つまり、神経細胞は最初から特定の方向に樹状突起を作ったり伸ばしたりするのではなく、トライアルアンドエラーを繰り返しながら、正しい方向に生えた樹状突起の一部を選択的に安定化させ複雑化させることによって、細胞全体として特定の方向に樹状突起を展開していくことがわかりました。
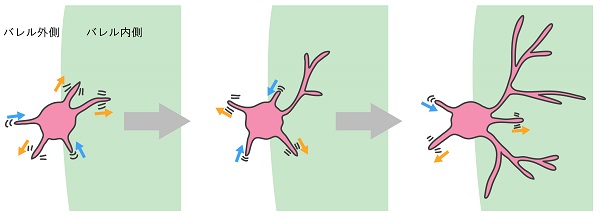
神経入力の空間的な偏りが樹状突起の競争を引き起こす
次に私たちは、神経入力が樹状突起の成長様式に与える影響を知りたいと考えました。新生仔期のバレルへの神経入力のソースは大きく2種類に分けられます。ひとつはヒゲを介して外部環境からの刺激により誘起されるもの。もうひとつは、ヒゲ刺激とは無関係に「自発的に」起こるものです。
自発神経活動は発達期の神経回路に広くみられる現象であり、回路の再編に重要な役割を果たしていると考えられています。最近私たちは新生仔期のマウス体性感覚野で、ヒゲが何かに触れたり動いたりしていないにも関わらず、あたかもヒゲに1本ずつ触ったかのようにバレルごとに発火するユニークなパターン(パッチワーク型)の自発神経活動を発見しました。この活動は視床皮質軸索を通じて体性感覚野へと届けられていました。
今回、ヒゲから脳へとつながる三叉神経を、ヒゲに近い部分で生後すぐに切断して体性感覚野へ入る自発神経活動がどうなるかを調べました。その結果、自発神経活動の頻度は正常マウスと比較してあまり変わらなかった一方で、その空間パターンは大きく異なっていました。つまり、このときバレルの神経細胞へはいろいろな方向から神経入力が入りうる状況になっていました。
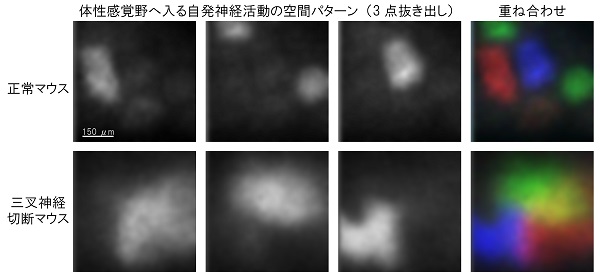
(上:正常マウス)通常時はバレルごとに発火するパッチワーク型の自発神経活動が見られる。このとき、バレルの神経細胞はバレル内側の樹状突起からのみ入力を受ける。
(下:三叉神経切断マウス)ヒゲから脳へとつながる三叉神経を切断すると自発活動の空間パターンが崩れた。このとき、バレルの神経細胞はあらゆる方向から入力を受けることができる。
そして、このマウスで生後3日目から6日目の長期イメージングを行ったところ、樹状突起のターンオーバー頻度が著しく低下しており、個々の樹状突起の形態は正常マウスでのバレルの縁の神経細胞のように、極端に複雑な一部の樹状突起や大部分を占める(生まれたばかりの)短い樹状突起は少なく、全体的に平均的な複雑さになることが明らかとなりました。もし、三叉神経切断によって神経入力の空間的偏りがなくなったことが樹状突起の成長様式に影響を与えたのであれば、正常なマウスでもバレルの中心に存在する神経細胞は三叉神経切断マウスの神経細胞と同様の挙動を示すことが予想されました。調べてみたところ、実際にそのようになっていることがわかりました。
バレルの縁にいる神経細胞はバレル内側に生えた樹状突起からのみ入力を受けることができますが、このとき、バレルの神経細胞は多くの樹状突起を作っては消すということを繰り返しながら、(おそらく)正しい相手とつながりを作った一部の樹状突起だけを勝者として成長させることにより、特異的な回路を形成すると考えられます。一方、三叉神経切断時やバレルの中心にある神経細胞はあらゆる方向から入力を受けることができます。このとき、樹状突起は競争する必要がなくなり、皆が“ほどほど”に成長すると考えられます。神経細胞が特定の方向からのみ適切な入力を受けられる時、すなわち入力の空間的偏りがある時、神経細胞は樹状突起の生存競争を誘導し、そのなかから適切な方向に生じた樹状突起を選択し安定化、成長させることにより、特異的な神経回路を形成するという仕組みが示唆されました。
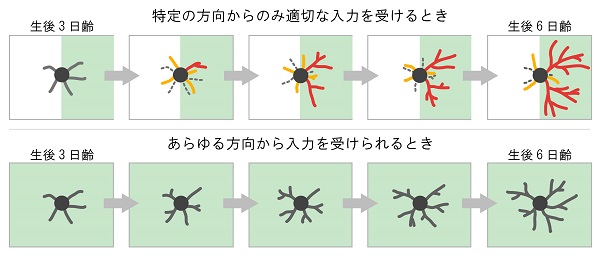
(下)入力に空間的偏りがない場合、樹状突起は競争する必要がなくなり、皆がほどほどに成長する
おわりに
今回、新生仔マウス脳の長期生体イメージングの成功により、新生仔期の回路再編機構の一端を明らかにすることができました。この成果を端緒として、回路再編過程での神経活動の経時変化や、そこにかかわる遺伝子の作用機序など、新生児期の神経回路再編機構の理解を今後さらに進めていきたいと考えています。
参考文献
この記事を書いた人

-
中沢信吾(写真左)
国立遺伝学研究所 形質遺伝研究部門 博士研究員。総研大生として同研究室にて学位取得。一つひとつの神経細胞がどのように作用し合い複雑な脳の機能を実現していくのかに興味があります。
岩里琢治(写真右)
国立遺伝学研究所 教授。総研大遺伝学専攻 教授(併任)。京大理院にて学位取得。マサチューセッツ工科大、理研BSIを経て2008年より現職。単に知識を増やすのではなく、新しい概念を生み出し、古典として長く残り多くの研究の礎となれる研究を目指しています。
ラボウェブサイト https://www.nig.ac.jp/labs/NeurGen/



