生命の起源 – 原始的な複製体はいかに複雑化しうるのか?
自己(情報)の複製と生命の起源
生命の起源を考えるためには、まず生命とは何なのかを考えないといけません。この哲学的な問いに対する決まった答えはまだありませんが、我々が生命だと疑いなく認識できるものには、少なからず共通の特徴があります。そのひとつが自己の複製、子孫を残すことです。細菌、植物、動物、いずれも子孫を残して繁栄します。そしてこの複製で重要なことは、遺伝情報(を担っているDNA)を複製し、後世に伝えることです。そのため生命の起源でも、遺伝情報の複製がキーワードのひとつとなっています。
これまでの研究から、生命の起源で遺伝情報を担っていたのは、RNAとよばれる4種類の核酸塩基(A/G/C/Uで表現される)が繋がったポリマー分子だったと考えられています(DNAの親戚みたいなもの)。またその4種類の塩基の並び順が情報となり、たとえば複製するといった機能を決定します。さて、原始地球において複製可能なRNAが生まれたとしましょう。ここから生命体へと近づいていくためにはどうすればいいでしょうか? 遺伝情報を増やしていって、たとえば代謝するなど、さまざまな機能を獲得し、複雑化していく必要があります。
先行した理論研究 – 複雑化は不安定
原始的なRNAの複製体はいかにして遺伝情報を拡張し、複雑化できるのでしょうか? この問いはまず1970年代、ノーベル賞受賞者のManfred Eigenによる一連の理論研究で脚光を浴びました。彼は原始生命のRNAは自身でたくさん情報を担うことはできず、情報の拡張には、異なる情報をもつRNA同士が協力し、複製する「ユニット」を形成することが必要だろうと示しました。これは、原始的な複製ではエラー(突然変異:たとえばAがGに変化する)が高頻度で起きると考えられており、長い一本のRNAにたくさんの情報を詰め込むと、複製でいずれかの情報が壊れる確率が高いからです。
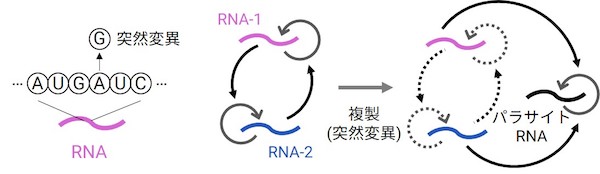
しかし、その後すぐに進化生物学の第一人者John Maynard Smithが大きな進化的問題を指摘しました。それは協力を搾取する利己的な複製体の出現に対して不安定であるということです。一般に、複製をすると、上述の突然変異が起きて、異なる特徴をもった個体が生まれることがあります(遺伝的多様性の創出)。ではもし機能を失う代わりに速く複製される、ウイルスのようなRNA(パラサイトRNAとよばれる)が出現するとどうなるでしょうか? 複雑な防御機構を持たない原始的な複製システムでは、パラサイトRNAばかりが複製され、機能をもつRNAはいなくなってしまいます(自然選択)。これは最も単純な進化の例です。さて、どうすればこのパラサイト問題を解決できるでしょうか?
この問いは50年も(!)理論的に研究され、どうやらパラサイトRNAを空間的に隔てる、細胞などの区画構造が必要そうだと示唆されていたものの、実験的な証拠が一切ありませんでした。誰も状況をうまく模擬する実験系を創れなかったのです。
実験研究へ – 協力的な複製システムの構築
そういうわけで私たちがやりました。実験系は(1)複数のRNAが協力的に複製する、(2)その際に突然変異が起き、進化可能である、(3)区画に封入されている、という3つの条件を満たす必要がありました。今回の研究ではこの構築が一番難しかったのですが(プロセスは複雑なので割愛します)、何とかできました。
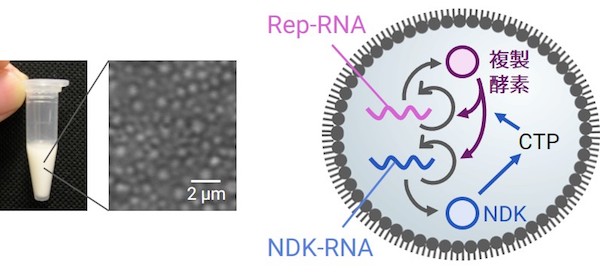
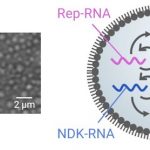
チューブの中に何十億もの区画構造があり(左)、それらに複製システム(右) が入っている。
油の中の水滴(エマルション)を細胞区画に見立て、その中にRep-RNAとNDK-RNAという2種類のRNAを入れました。それぞれは複製酵素と核酸塩基の合成酵素(NDK)という異なる遺伝情報をもっています。またこの情報を読み出す(翻訳する)ための機械を細菌から借りて一緒に封入しています。すると、まずNDKがNDK-RNAから翻訳され、それが核酸塩基のひとつであるCTP(A/G/C/UのうちC)を合成します(代謝の一種)。そして複製酵素(Replicase)がRep-RNAから翻訳され、そのCTPと、最初から入れておいたA/G/Uを繋げてRNAを複製します。つまり、互いの複製に互いが必要な、協力的な複製システムになっています。なお、このシステムは、RNA・プロテインワールドとよばれるDNAが誕生する前の時代の、原始細胞を想定したモデルとなっています。
ScyllaとCharybdisの狭間で生き延びる
次に本システムを用いて、試験管内で進化を模擬した実験を行い、協力的複製が長期的に維持されるかを調べました。進化実験では、(1)複製反応、(2)希釈、(3)区画の融合分裂(原始的にありえた細胞分裂を模擬)というサイクルを繰り返しました。複製の際に突然変異が入り、また十分に複製できないと希釈によって淘汰されます。さて、まず協力RNAを高濃度にして複製をスタートした場合は、すぐにパラサイトRNAが大量に出現して協力RNA(Rep-RNAとNDK-RNA)の複製は止まり、希釈されてしまいました。また逆に、低濃度から複製をスタートすると、今度は2種類の協力体は同じ区画内で共存できず、複製できませんでした。
この現象は理論的に予測されており、その差し掛かると死を意味する両方向の脅威から、生命の起源のScyllaとCharybdis(ギリシャ神話の怪物)とよばれています。しかし一方で、興味深いことに、パラサイトRNAの出現を抑制しつつ協力的複製が可能な、中間の濃度領域の存在を見い出しました。しかも驚くべきことに、一度この領域で複製が開始されてしまえば、なんと協力関係は低濃度で安定化し、大きな希釈が起きても維持されるようになっていました。この安定化はこれまでの理論研究でも知られていなかった現象であり、コンピュータシミュレーションの結果、どうやら区画の融合分裂頻度に依存して起きるようだとわかりました。
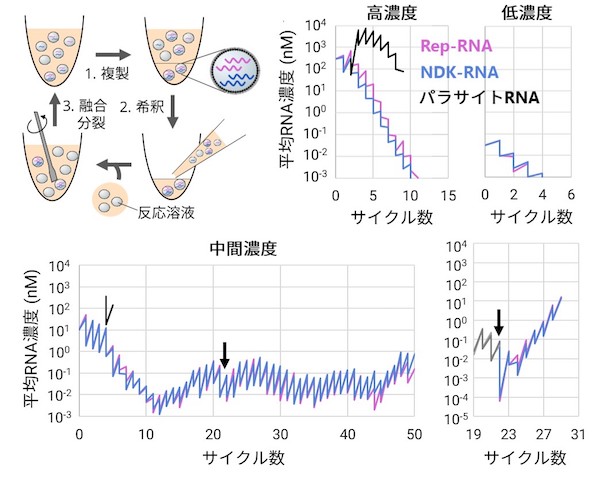
サイクル実験(左上)を高濃度または低濃度で開始すると複製は維持されないが、維持される中間濃度領域が存在する。この領域では大きな希釈にも耐えられる(矢印、右下)。
協力的なRNA複製体の進化
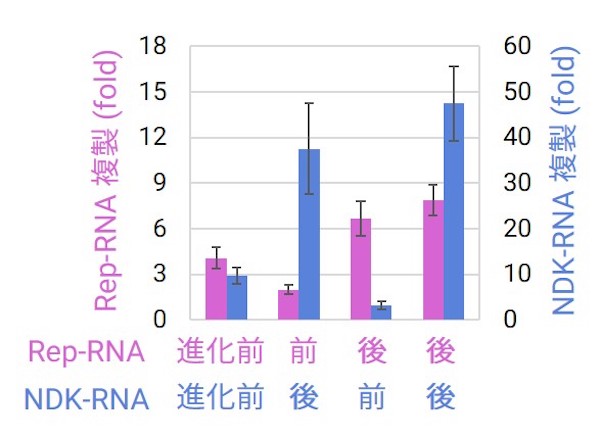
突然変異は時に機能の低下や欠損をもたらしますが、また別の時には機能を高めたり、機能を維持したまま複製を速くするなど、有益にはたらくこともあります。そこで長期の複製を経た後のRep-RNAとNDK-RNAに挿入された変異の効果を調べてみました。すると、最もよく見られた変異セットをもつペア(進化後)は、元々のペア(進化前)と比べて高い協力的複製を示しました。これは変異が有益で、協力体が進化したことを示しています。
では一方で、進化後のRNAと進化前のRNAを組み合わせてみるとどうでしょうか? なんと進化後が進化前のパートナーの複製を阻害してしまいました。詳しく調べると、進化後の協力RNAはいずれも複製が速くなっており、進化前のパートナーRNAからすると利己的な複製体になっていました。ところが進化後同士のペアはうまく協力できるのです。この一見矛盾するように聞こえる結果は、個々の協力RNAの利己的な進化と、それに続くお互いへの適応的な進化により説明することが可能です。この結果は今まで知られていなかった、新たな進化のメカニズムの存在を示唆しています。
おわりに
本研究では、原始的な複製体の複雑化における問題点と、それが解決可能であること、さらには複製を続けてよりよい複製体へ進化もできることを実験的に示しました。でももちろん、これは生命の起源という壮大な謎のほんの一片の理解に貢献したにすぎません。最初のRNAや区画構造がどうやって生まれたのか、それらの材料はどこから来たのか、どうやって複製を開始したのか、解決すべき問題は山積みです。でもそれはもう、時間の問題かもしれません。この分野の大御所であるJohn Sutherlandの昨年の論文のタイトルはこうです。 “Studies on the origin of life — the end of the beginning”「生命の起源研究 ― 始まりの終わり」。そう、準備は整ったのです。生命の起源研究のこれからの急展開をご期待ください。
参考文献
- Ryo Mizuuchi & Norikazu Ichihashi (2018). Sustainable replication and coevolution of cooperative RNAs in an artificial cell-like system. Nature Ecology & Evolution, 2, 1654–1660.
- Ricard Solé (2018). Cooperation in an RNA world. Nature Ecology & Evolution, 2, 1527–1528.
- John D. Sutherland (2017). Studies on the origin of life—the end of the beginning. Nature Review Chemistry, 1, 0012.
この記事を書いた人

-
日本学術振興会海外特別研究員(ポートランド州立大学)
2018年、大阪大学大学院情報科学研究科博士課程修了。在学中にNASA Ames研究所での長期インターンシップ、日本学術振興会特別研究員(DC1)を経て2018年4月より現職。2016年より慶応大学Advanced Astrobiology Projectの外部アドバイザーを務める。専門は生命の起源と初期進化だが、奄美大島のフグのミステリーサークル(産卵床)も研究している。
この投稿者の最近の記事
 研究成果2018年10月1日生命の起源 – 原始的な複製体はいかに複雑化しうるのか?
研究成果2018年10月1日生命の起源 – 原始的な複製体はいかに複雑化しうるのか?


