深海熱水噴出域から浅海へ旅する貝の幼生- 海洋表層で分散するミョウジンシンカイフネアマガイ
海底火山や海嶺、海洋プレートの沈み込み帯周辺には100℃を超える海水が吹き出す熱水噴出域があり、固有の貝やエビ・カニ・ゴカイ類が生息しています。これらの動物は、太陽光エネルギーに由来する光合成生産には依存せず、熱水に含まれる硫化水素やメタンを用いたバクテリアの化学合成生産に支えられており、深海としては例外的に高密度の集団を形成します。一方、数十年から数百年の時間スケールでみると、個々の噴出域における熱水活動は、衰退・停止することがあります。よって、熱水性種が個体群を維持するためには、離れ小島のように分布する熱水噴出域間を何らかの方法で移動することが必須となります。
本稿では、深海熱水噴出域に生息する貝類の一種が、プランクトン幼生期に太陽光の届く水深帯まで鉛直遊泳し、表層流にのって遠方の熱水噴出域へ移動するという初の実験的証拠について紹介したいと思います。

堆積物表面には白色のバクテリアマット、ミョウジンシンカイフネアマガイ(中央上部)とフジツボ類の一種(右上部)の群集がみられる。海洋研究開発機構の海洋調査船「なつしま」ならびに無人探査機「ハイパードルフィン」により撮影(NT14-06航海、第1652潜航) (c) JAMSTEC
幼生分散の役割
熱水噴出域に生息する無脊椎動物の多くは、成体の移動能力が乏しい一方で、卵や幼生として海中を浮遊する特徴をもちます。この浮遊期における海洋分散が種の分布域を規定し、また絶滅を防ぐために重要な役割を果たします。
熱水性動物の幼生発達様式は、母親から受け取った卵黄を栄養源として成長する卵黄栄養型と、他の浮遊生物を食べて成長するプランクトン栄養型に二分できます。前者の幼生は、海底近くでの採集例があり、また、低水温・高圧力の実験環境下においてよく成長するため、底層流によって分散すると考えられています。一方、後者のプランクトン栄養幼生は、孵化後に中深層へ移動し、摂餌・成長すると推測されてきましたが、幼生の実際の行動や分散水深についてはほとんど解明されていませんでした。
プランクトン栄養幼生の分散水深を探る
今回、私は狩野泰則准教授・小島茂明教授(東京大学大気海洋研究所)および渡部裕美博士(海洋研究開発機構)と共同で研究を行い、沖縄トラフおよび伊豆・小笠原の熱水噴出域(水深442〜1227m)から、プランクトン栄養型の貝類であるミョウジンシンカイフネアマガイの成体と卵嚢を採集し、幼生飼育および遺伝子配列の比較により同種の分散機構を検討しました。
シンカイフネアマガイ類は、1)太平洋・大西洋・インド洋の熱水域に広く分布する点、2)大気圧下で成体と幼生の飼育が可能であり、卵嚢が透明で内部の胚発生を追える点、3)貝殻形態から発生様式および幼生の着底サイズを推定できる点で、生態情報に基づいた幼生分散の研究対象として優れた分類群です。試料の採集は、海洋研究開発機構の海洋調査船「なつしま」と無人探査機「ハイパードルフィン」を用いて実施しました。
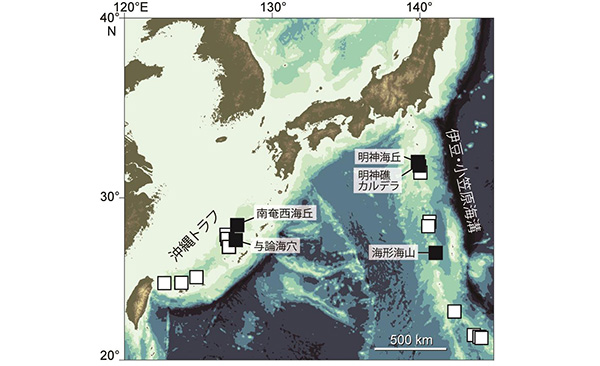
黒塗りはミョウジンシンカイフネアマガイ成体の生息地点で、地点間は最大1300km離れている。
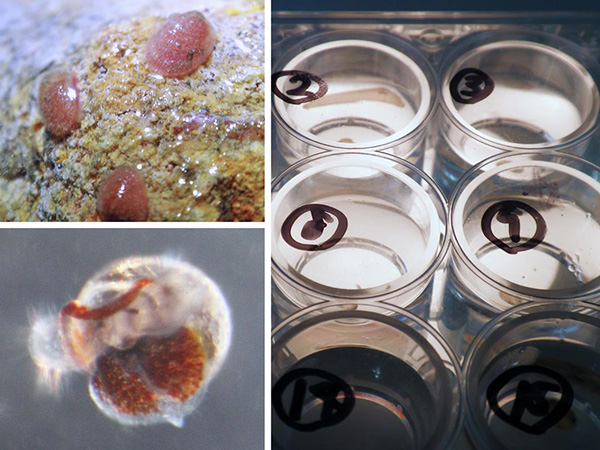
右:同種孵化幼生の飼育実験
5℃・10℃・15℃・20℃・25℃・30℃条件において各10個体を1個体ずつ個別に6か月間飼育し、成長と生残の至適水温を評価した。
左下:25℃給餌条件の孵化後112日幼生(殻長260µm)
ミョウジンシンカイフネアマガイの海洋表層分散
卵嚢から孵化した幼生を実験室で観察したところ、すべての個体が継続的な上昇遊泳を行い、その速度は5℃・25℃の条件下で毎分16.6〜44.2mmに及ぶことがわかりました。また、これらの個体は、無給餌条件下でも5℃・15℃・25℃で各平均156・84・30日間を生きながらえました。つまり、水深1000mを超える深海環境から、幼生が餌なしで表層まで到達できることを示唆します。
植物プランクトンである珪藻の一種を給餌した180日間の幼生飼育実験では、成体の生息環境に近い10℃ないし15℃においてほぼ無成長のまま死亡する一方、分布域の表層水温に近い25℃で最高の生残・成長率を示すという、幼生の鉛直行動と整合的な結果が得られました。また、幼生が本種の着底サイズ(殻径720µm)に至るには、この25℃条件の成長率で1年以上の期間を要すると推定されました。そこで、地理的分布を網羅するよう採集した77個体の成体を用いてミトコンドリアDNA配列を比較したところ、最大で1300kmほど離れた生息地間を頻繁に行き来していることが認められ、長期にわたる浮遊幼生期の存在が支持されました。
以上の結果は、ミョウジンシンカイフネアマガイ幼生が深海熱水噴出域で孵化した後に太陽光の届く水深(200m以浅)まで泳いで浮上、植物プランクトンを食べて成長し、表層海流にのって長距離分散することを強く示唆します。
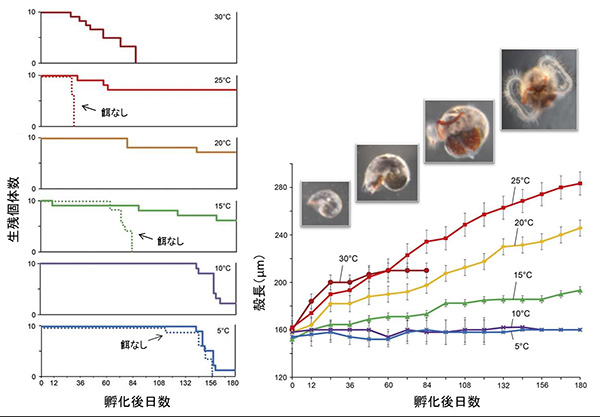
実線が給餌、点線は無給餌条件を示す。同種地理的分布域の表層水温に近似する25℃の給餌条件で、最も良い成長・生残がみられた。成体の生息環境水温(11℃前後)では、ほとんど成長せず死亡する。30℃の高温でも長期間生存できない。
表層水温と熱水性動物の地理分布
ミョウジンシンカイフネアマガイの成体は、伊豆・小笠原海溝と沖縄トラフ北部の熱水域のみから報告され、その南に位置するマリアナ海域や沖縄トラフ南部の熱水環境からは知られていません。これら南方の海域では、夏季の平均表層水温が29℃を超えるため、幼生は長期間生存できずに死亡する(生残至適水温を参照)と考えられます。つまり、表層水温が、はるか深くに生息する熱水性動物の分布を規定している可能性があります。
興味深いことに、北極や南極の熱水噴出域では、生息するすべての動物が卵黄栄養型の発生様式を示します。これら極域の熱水生物群集は、直上の表層水温が年間を通して−1℃から2℃と低く、また餌となる植物プランクトンが僅かな時季にしか得られないため、長期の浮遊期を経て着底するプランクトン栄養型発生に不適であると考えられます。このことからも、表層環境が深海熱水動物の分布を規定している可能性を推察できます。
幼生はどのように熱水域を見つけるのか
生まれた場所を離れた幼生は、表層で成長した後にどのように熱水噴出域を見つけるのでしょうか。未だはっきりしたことはわかっていませんが、幼生は熱水に由来する水温変化や特徴的な化学物質を認識し着底すると考えられます。また、分散の最中に捕食されるなど、熱水域に辿りつくことなく死亡する幼生も数多く存在すると推測されます。どれくらいの幼生の放出・供給によって個体群が成り立っているかについても今後の研究が待たれます。
今回の研究は、深海熱水噴出域に生息する固有種の一部が、プランクトン幼生期に表層まで遊泳し、長距離分散することを明らかにしました。また、表層水温が、深海に生息する熱水域動物の地理的分布を規定している可能性が示されました。これは、化学合成群集の生態・進化に関わる新たな仮説であり、熱水噴出域と光合成環境の物質循環にも新たな視点を加えるものと考えられます。
参考文献
Yahagi, T., Kayama Watanabe, H., Kojima, S. & Kano, Y. Do larvae from deep-sea hydrothermal vents disperse in surface waters? Ecology 98: 1524–1534 (2017). DOI:10.1002/ecy.1800
Yahagi, T., Kayama Watanabe, H., Kojima, S. & Kano, Y. Larval connectivity between deep-sea hydrothermal vents via surface waters. The Bulletin of the Ecological Society of America 98: 231–235 (2017). DOI:10.1002/bes2.1324
Yahagi, T., Fukumori, H., Warén, A. & Kano, Y. Population connectivity of hydrothermal-vent limpets along the northern Mid-Atlantic Ridge (Gastropoda: Neritimorpha: Phenacolepadidae). Journal of the Marine Biological Association of the United Kingdom, in press.
この記事を書いた人

- 東京大学大気海洋研究所 特任研究員。2017年3月 東京大学大学院新領域創成科学研究科博士課程修了、博士(環境学)。専門は海洋生態学。深海熱水噴出域における底生生物群集の成立過程や極限環境への適応機構に興味をもっています。学術研究船・探査機を用いたフィールド調査をはじめ、国内外の熱水噴出域で採集された生物を対象に、分類、飼育、遺伝学、分析化学手法を用いて多角的に研究に取り組んでいます。