外来種のモウソウチク・マダケが里山生態系を脅かす – 温暖化が進めば北日本でも分布拡大する可能性
日本文化と竹林
イネ科タケ亜科(87属1500種以上)は世界中の熱帯から温帯に分布し、日本にも150〜600種が分布するとされています。このなかで、日本で高さが5mを超えるような竹林を形成するのはマダケ属のタケ(マダケ類:モウソウチク(孟宗竹)、マダケ(真竹)、ハチク(淡竹)、クロチク(黒竹)、ホテイチク(布袋竹)、キッコウチク(亀甲竹))です。林野庁の2012年の最新統計では、日本の竹林は16万1千ヘクタールに上り、その99%をモウソウチクとマダケが3:1の割合で占めています。
マダケ類は日本でも食料(主にモウソウチクとハチク)や材(主にマダケ)、園芸などの目的で利用されてきました。マダケ類は日本文化に古くから関わってきたため、これらが外来種であるというと驚かれるかもしれません。しかし、マダケ属の自然分布域は中国の長江流域以南で、モウソウチクは18世紀前半に、他のマダケ類は8世紀頃に中国から持ち込まれた外来種だと考えられています(最近になって神奈川県相模原市の200〜300万年前の地層からマダケあるいはその近縁種の化石が発見されたので、日本のマダケが当時からずっと生き残ってきたのか、それともメタセコイアやイチョウのようにある時点で絶滅したのか、今後の研究が待たれます)。
多くの日本人に親しみ深い竹取物語が成立した9世紀後半から10世紀前半にはモウソウチクはまだ移入されていなかったので、かぐや姫(三寸ばかりなる人)が入ることができる竹はマダケかハチクだと考えられています。当時まだエキゾチックで貴族だけが楽しむことができた竹林に、エキゾチックなかぐや姫のイメージを重ねたのだと考える人もいます(まさにexotic species=外来種ということになりますね)。

放棄竹林の増加
しかし1970年代以降にはタケノコの輸入自由化や農家の高齢化に伴って竹林の管理放棄が進みました。特に成長の早いモウソウチクは、タケノコから約1か月で20mもの高さに成長し、 周りの植物を日陰にして枯らしてしまいます。放棄竹林は年に最大3〜4mの速さで周囲に拡大していくことから、里地里山管理のうえで最大の問題点とも言われています。2015年には、国がマダケ属のタケを産業管理外来種(産業または公益的役割において重要であるが、利用上の留意事項が求められるもの)に指定しました。各地で駆除や有効活用が試みられていますが、再生の速い竹林を完全に駆除する多大な労力を要することもあり、根本的な解決には至っていません。
前述のようにマダケ類は南方種なので、放棄竹林は主に西日本で問題になっていますが、現在マダケは青森県、モウソウチクは北海道南部まで分布しています。将来の気候変動に伴い日本域の温暖化が進めば、さらに高緯度・高標高でも竹林が生育可能となり、地域の生態系を乱してしまう可能性があります。
一方、現在の日本の竹林の分布は主に人間によって形成されたと考えられます。このため、マダケ類が生育可能な地域が今後どのように拡大していくのかを前もって知ることは、将来の竹林の管理を考えるうえで重要です。
マダケ・モウソウチクの分布予測
将来のタケの分布を予測するため、私たちは長野県から青森県までの145か所のAMeDASから半径5km以内のマダケ・モウソウチクの有無を調べました。さらに、1975〜1980年の空中写真から過去の竹林分布も調べ、この期間に新たに竹林が定着した場所が17か所あることを見つけました。
これらのデータと緯度・標高との関係を見ると、1975年以降に新たに成立した竹林は、緯度-標高傾度に沿った分布限界付近に集中していることが明らかになりました。このことは、過去40年程のあいだにもマダケ類が植栽され、竹林の分布域が拡大していたことを示唆しています。
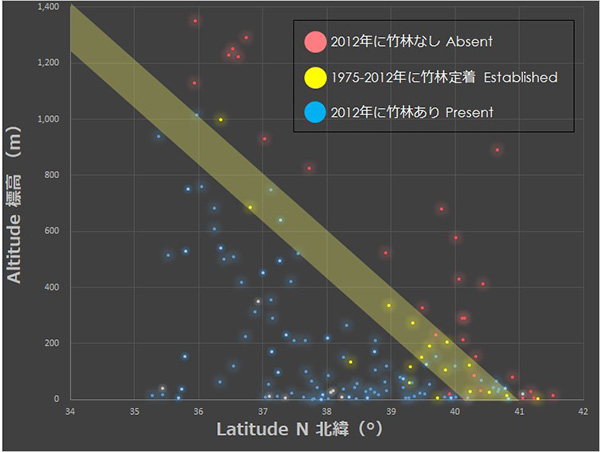
さらに私たちは調査で得られたデータとAMeDASから得られた2002〜2010年の気候の平均値を結びつけ、モウソウチク・マダケの分布を予測する生態学的ニッチモデルを作成しました。
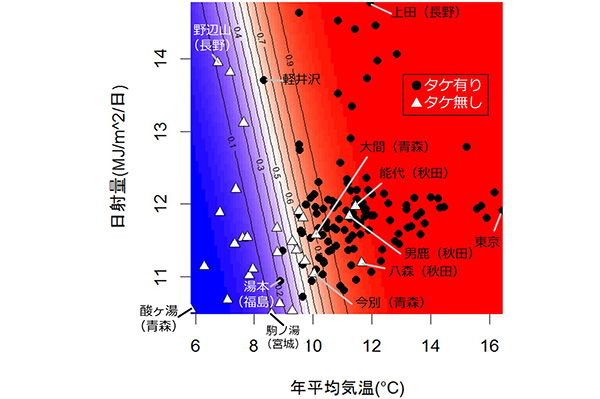
その結果、平均気温と日射量の年平均を説明変数としたロジスティック回帰モデルによって145か所の竹林有無を93%の正答率で予測することができました。このモデルでは年平均気温が8〜11°Cを超えるとモウソウチク・マダケが生育できる確率が高くなり、日射量も正の効果を持っていました。しかし確率が高い場所でも、竹林が確認できなかった地点(能代、男鹿、八森、大間など)が見られました。これらの場所は、気候条件的にはモウソウチク・マダケが生育できますが、まだ植栽が行われていない場所と考えられます。
将来のタケの生育可能域の予測
将来のタケの生育可能な地域を知るためには将来の気候、気温と日射量の値を予測する必要がありますが、このためには大気大循環モデルという気候モデルが使われます。将来気候を決定するためには二酸化炭素を始めとする温室効果ガスの濃度を指定する必要がありますが、これは排出シナリオと呼ばれる将来予測に基づいて決定されます。温室効果ガスによる人為的な気候への影響を少しでも緩和しようとする取り組みが各国・世界規模で行われていますが、それがどれだけ奏功するかは今後の経済発展・技術進歩、さらには人々の環境意識などに依存しています。研究者としては可能性のある複数の排出シナリオを用いて、可能な限り信頼性の高い予測を行うことが求められています。
本研究で用いられた年平均の気温や日射量といった気候の将来変化は気候モデルによって予測されますが、気候モデルは完全ではないために「予測の誤差」(バイアス)が存在することが知られています。バイアスは現在と将来で大きくは変わらないと考えられますので、現在気候と将来気候の「差」はかなりの信頼度で予想することが可能なのですが、タケの生育可能地域を考える際には実際の気温や日射量の値が必要となります。
そのため本研究ではAMeDASデータと現在気候のモデル最現値を比較することでバイアスを推定し、この分だけを将来気候から差し引くというバイアス補正によって将来気候を推定しました。
前述のようにタケの生育可能性を決定するうえで最も重要な要素は気温ですが、気温の将来変化は場所ごとに異なるため、個々の地点での気候変化をできるだけ空間詳細な情報として出すことが必要です。そのために用いられるダウンスケーリングと呼ばれる手法では、最初に地球全体の気候を再現する大気大循環モデルを動かしておおまかな気候を再現し、その結果を基に日本付近で空間解像度の高い領域気候モデルを動かします。私たちの研究では全球平均した気温が産業革命前から4℃昇温したという設定のもと、日本域で水平方向に5kmのグリッドで将来気候を再現しました。

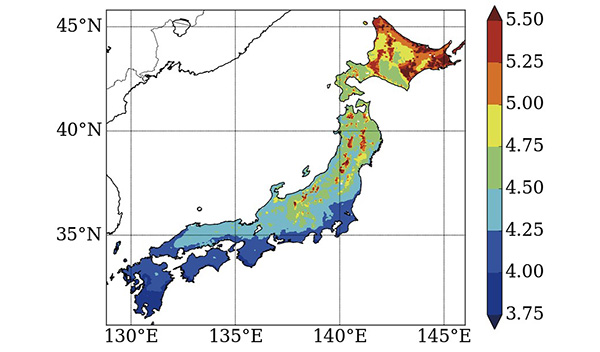
図はこの結果から得られた昇温量ですが、値が場所ごとに大きく異なっていることがわかります。全体的に見ると北日本で大きな昇温が得られており、特にオホーツク海沿岸でその傾向が顕著に見られます。これは将来の温暖化の影響は全球規模で゙見て熱帯域よりも高緯度域で顕著に表れやすいこと、さらにオホーツク海沿岸では冬季の流氷が減少することで気温が大きく上昇することが原因と考えられています。
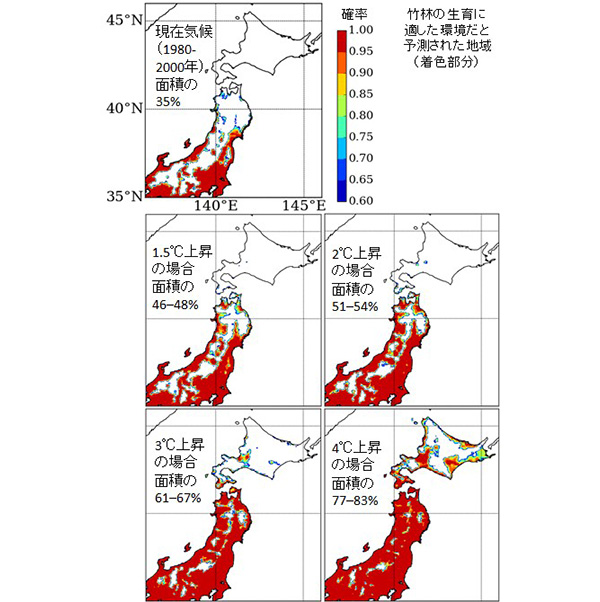
このようにして得られた気温と日射量の将来での値を竹林生態モデルに入れることで、4℃昇温した世界での日本域のタケの生育可能な地域を求めました。2015年のパリ協定によって産業革命前から全球昇温を2℃未満に抑えることが目標とされ、さらに努力目標として1.5℃未満の昇温量が設定されました。これら温暖化進行のさまざまなレべルのもとでの影響を評価するために、 前述の4℃に加えて1.5℃、2℃、3℃といったそれぞれの昇温量のもとで竹林の生育可能域を推定しました。

その結果、1980〜2000年には東日本(北緯35度以北、東経136度以東:図の範囲)でモウソウチクとマダケの生育に適した土地の割合は35%であったのに対し、日本の平均気温が産業革命前に比べて1.5℃上昇した場合には46〜48%、2℃上昇では51〜54%、3℃上昇では61〜67%、4.0℃上昇した場合には77〜83%まで増加し、北限は稚内に到達すると予測されました。
まとめ
これらの結果は、地球温暖化を1.5℃に抑制するパリ協定の目標が、温暖な生育環境を好む外来種の分布拡大を抑制するために、一定の効果を持っていることを示しています。一方で、タケの広域の分布は主に人間による植栽によって決まっています。今回私たちが予測したのはタケの「潜在」生息適域であるため、分布確率が高い場所でも人間が植えない限り、タケが生えることはありません。温暖化がある程度進んでしまった場合にも、外来種被害予防三原則である、入れない・捨てない(管理放棄しない)・拡げない(タケを新たな土地に定着させない)といった管理と対策を、地域住民と行政が一体となって進めることが重要です。
このように、気候変動対策においては、温暖化そのものを抑制する温室効果ガスの削減(緩和策)と、気候変動が進んでしまった場合の適応策の両方を進めていく必要があります。
参考文献
Takano KT, Hibino K, Numata A, Oguro M, Aiba M, Shiogama H, Takayabu I, Nakashizuka T. 2017. Detecting latitudinal and altitudinal expansion of invasive bamboo Phyllostachys edulis and Phyllostachys bambusoides (Poaceae) in Japan to project potential habitats under 1.5°C– 4.0°C global warming. Ecology and Evolution. DOI: 10.1002/ece3.3471
国立科学博物館 筑波実験植物園 植物化石展ブログ 化石の見方~高校生たちの取り組み
環境省. 2004. 里地里山パンフレット~古くて新しい いちばん近くにある自然~ p.4 竹林が里 地里山を飲みこむ.
この記事を書いた人

-
髙野(竹中)宏平 (写真左)
長野県環境保全研究所 自然環境部 技師・植物生態担当(地球環境科学博士)。千葉県松戸市生まれ。東京農工大学農学部を卒業後、北海道大学大学院地球環境科学研究科に進学し、オオバナノエンレイソウの自殖性の進化(修士)、タロイモショウジョウバエとサトイモ科植物の送粉共生(博士)などを研究。長崎大学熱帯医学研究所(ベトナム産マラリア媒介蚊の分子系統、ロタウイルス外皮タンパクの分子進化)、総合地球環境学研究所(ボルネオの熱帯雨林の生物多様性)、東北大学(気候変動の生態系影響)、森林総合研究所(長野県の生態系の気候変動適応)を経て2017年4月より現職。

日比野研志(写真中央)
東京大学生産技術研究所特別研究員。京都大学理学研究科卒業(理学博士)。筑波大学研究員を経て2016年より現職。これまではさまざまな時空間スケールでの気候変動の影響評価を行ってきました。特に降水などの気候値の将来変化の定量評価やそれらの不確実性の考察、小スケールでは積乱雲や台風に伴う極端降水の将来変化を、気候モデルのアンサンブル実験などによって研究してきました。現在は、陸面モデルや河川モデルなども使い、全球的な水循環の理解や 洪水・干ばつなどの予報技術にも関心を持っています。
小黒芳生(写真右)
森林総合研究所特別研究員。東北大学生命科学研究科卒業。生命科学博士。東北大学産学官連携研究員を経て、2016年より現職。これまで花の化学的性質を介した植物と花を食べる動物との相互作用の研究、樹木の病気の空間分布を予測する研究などを行ってきました。自分は生きものの相互作用や進化を研究する人だと思っていたのですが、気がついたらデータ解析・処理を 相談される人になっていました。現在はさまざまなデータを結びつけて生態系サービス(自然の恵み)を予測、評価、地図化する研究を行っています。



