コオロギ求愛ソングにおけるピッチ安定性 – 音痴な鳴き声ではメスに振り向かれない?
私たちは普段、さまざまな方法でお互いにコミュニケーションをとり、社会生活を送っています。 人間以外の多くの動物も、音(空気の振動)を耳や全身で感じ取ることによって、餌をとったり、天敵から逃れたり、あるいは求愛行動を行ったりして生活しています。これはイヌやウサギのような脊椎動物から、セミやコオロギのような昆虫(無脊椎動物)までよく共通した特長と言えます。
人間以外の動物は、人間のように高度な文法を使いこなすことはできませんが、 それぞれに 固有の「鳴き声パターン」を使い分けることによってお互いにコミュニケーションをとっていると考えられています。「鳴き声パターン 」はいわば「ことば」 のようなもので、動物の「ことば」を解読することが広く試みられています。動物の「ことば」を解読することができれば、動物たちのコミュニケーションをより深く理解することができ、より複雑な自然現象を解き明かす手がかりとなります。
コオロギが声を発する仕組み
私は動物の鳴き声によるコミュニケーションについて調べるために、コオロギ(和名:フタホシコオロギ、学名:Gryllus bimaculatus)を使って研究を行なってきました。本当はセミのほうが面白いと思ったこともあるのですが、セミは幼虫の期間が何年も続いた後、成虫となって鳴き声を発する期間が数週間しかない(それに飼育も難しい)ということから、私の現在の境遇では取り組むことが難しい研究対象であると考え、断念しました。
脇道に逸れましたが、コオロギが「声」を発する仕組みは、私たちが声を発する仕組みとはだいぶ違います。私たちは喉の奥にある声帯を振動させ、さらにそれによって生み出された空気の振動を気道で共鳴させて発生していると考えられます(さらに口の開き方や舌・口唇の使い方などが最終的な声を決めますが、詳細はここでは割愛します)。
一方、コオロギの場合には前翅を振るわせることによって空気の振動を生み出し、鳴き声を発していると考えられています。また、コオロギの場合、翅をこすりあわせることによって音を出すことができるのは、通常オスのみです。これは、鳴き声を奏でるのに必要な「つめ」と「ヤスリ」の構造が、オスの翅にしかないためです。
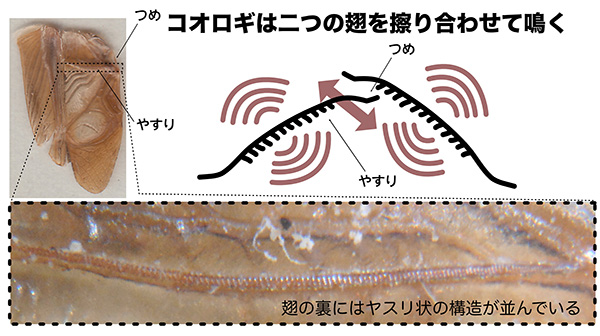
左上写真はフタホシコオロギ(オス)の前翅。翅の裏にはヤスリ状の構造(下拡大写真)があり、片方の翅の「つめ」をもう片方の翅の「やすり」にこすりつけることにより、翅全体を振動させて音を発する。
コオロギの鳴き声とメスの「選り好み」
コオロギのオスはいくつかの鳴き声パターンを持っていますが、私は特にメスを交尾へと誘うための2種類の鳴き声に着目して研究を行なってきました。オスは「呼び鳴き(calling song)」とよばれる鳴き声によって、遠くにいるメスを近くへと引き寄せ、近づいてきたメスを「口説き鳴き(courtship song)」と呼ばれる鳴き声によって交尾へと誘います。
一般的に(カブトムシなど)虫の交尾ではオスがメスの背中にマウントする形で交尾を行いますが、コオロギの場合は 、メスがオスの背中にマウントするという特殊な形式で交尾が行われます。この理由はよくわかっていませんが、ひたすら鳴き声を奏でてメスを待ち続けるコオロギの姿は、とても紳士です。また、この特徴によって、コオロギを用いると「メスによるオスの選り好み」を比較的簡便に観察することができます。
たくさんのオスを観察していると、あるオスの鳴き声には人気・不人気があることがわかります。つまり、あるオスの鳴き声にはメスがよく口説かれる一方で、別のオスの鳴き声にはまったくメスが振り向かない、ということがしばしば見受けられます。しかし、1)オスの鳴き声にどのような個性があり、2)どのような鳴き声がメスに好まれるのか、ということについては、いくつかの断片的な研究のほかにわかっていることはあまり多くありません。
ひとつの仮説としては、メスがより体サイズの大きいオスを好むということが考えられます。体の大きいオスは、ある環境により適応した個体であると考えられるからです。そこで私は今回、鳴き声のパターンと体サイズとの関連を調べることにしました。
どんな大きさの個体でも、鳴き声の高さは一定
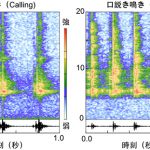
鳴き声のパターンを研究するためには、鳴き声の「音色」を研究者(人間)が理解できるように可視化することが必要です。そのようなテクニックとしてスペクトログラムという技術がよく用いられます。これは、鳴き声に含まれる周波数成分(音の高さに相当するパラメータ)を色で表示するもので、波形データと合わせることで、鳴き声のパターンを視覚的にわかりやすく表示することができます。
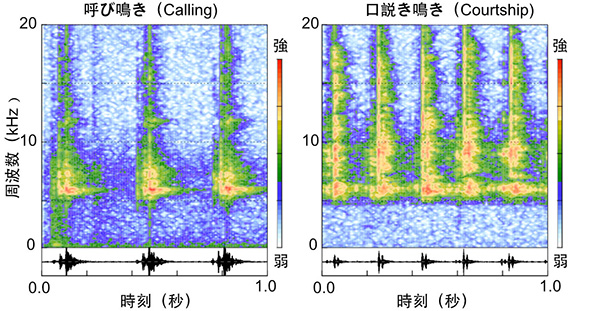
それぞれ1秒間の呼び鳴き(左)および口説き鳴き(右)のスペクトログラム。横軸は時刻、縦軸は周波数帯を示し、赤色ほど強度が高いことを示す。呼び鳴きは5.8kHz付近に単一周波数ピークを示し、口説き鳴きは5.8 kHz付近のピークに加えて、高音成分(~18kHz)に広く周波数成分を有し、さらに鳴きのリズムも早い。それぞれスペクトログラムの下部には波形を示してある。図で示したのは羽化後2週目の成熟した個体から録音した鳴き声。
このスペクトログラムに着目することによって、これまでにいくつかのことがわかってきました。まず、呼び鳴きと口説き鳴きでは、それぞれ特有の周波数ピークが存在します。これらの周波数の値(つまり音の高さ)は、コオロギの翅にあるミラーやハープと呼ばれる領域の振動数と密接に関連していると考えられていますが、最近私は、フタホシコオロギのこれらの鳴き声について、体全体の大きさやミラー部・ハープ部の大きさに影響されずに、常に一定の周波数になるように調節されているということを見出しました。
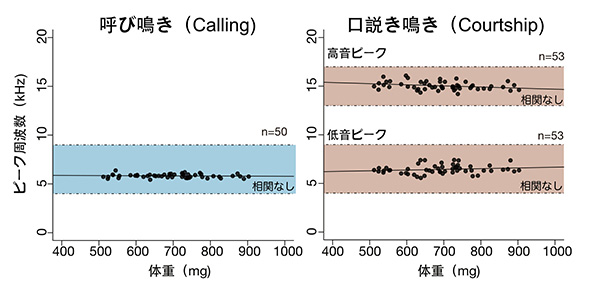
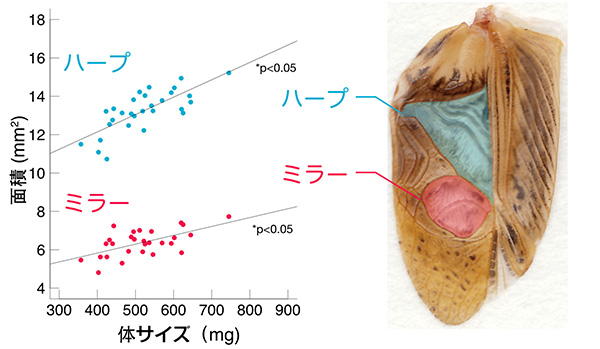
実際この結果は、ほかの研究者たちによって提唱されている、メスの耳は呼び鳴きの周波数に対してチューニングされており、特に音源定位(音がどの方向から来たかを認識すること)の感度が呼び鳴きのピーク周波数周辺で高くなっているという仮説ともよく合致しています。これらのことを踏まえると、フタホシコオロギにとっては鳴き声の高さが非常に重要な意味を持っており、ある一定の音程から外れた鳴き声を発するオスは、メスに見向きもされないということが推察されます。
残された疑問と今後の展開
これまで、コオロギの発声においては、つめとヤスリで生じた振動がミラーやハープ「共鳴(共振)」によって増幅され、最終的な鳴き声として発せられていると考えられてきました。ミラーやハープのような膜状構造体の振動特性は、形状・素材・張力といったさまざまな要因によって決定されますが、一般的にはサイズが大きくなるほど固有振動数の値は小さくなると考えられます。
したがって、今回の発見はコオロギの鳴き声の高さが翅のサイズ以外の要因(張力の調節など)によって、最終的な音の高さが一定になるように厳密に制御されていることを示しています。 また神経科学的には、コオロギが自分の鳴き声を耳で聞き、フィードバック学習を行なっている可能性が考えられます。さらに極端な仮説としては、ミラーやハープは実際には「共振」しておらず、つめとヤスリで生じた振動がそのまま「強制振動」を引き起こしている可能性も考えられます。ただしその場合には、どのようにして大音量を維持しているのかという別の疑問も生じます。
このように、一見単純に見えるコオロギの鳴き声ですが、まだまだ多くの謎が隠されています。私は今後まず、鳴き声のパターンの成熟過程に着目し、成長に伴って上記の鳴き声パターンがどのような過程をたどって獲得されるのかを明らかにしようと考えています。さらに最終的な目標としては、周波数以外のさまざまなパターンも含めて、メスがどのような鳴き声を好むのかを明らかにしようと考えています。
この目標に向けて、現在は行動実験や計算機を活用した解析システムの樹立を試みています。新しい研究成果が得られ次第、またご報告できればと思います。
参考文献
1. Miyashita, A., Kizaki, H., Sekimizu, K. & Kaito, C. No Effect of Body Size on the Frequency of Calling and Courtship Song in the Two-Spotted Cricket, Gryllus bimaculatus. PLoS One 11, e0146999 (2016).
2. Anichini, M., Kuchenreuther, S. & Lehmann, G. U. C. C. Allometry of male sound-producing structures indicates sexual selection on wing size and stridulatory teeth density in a bushcricket. J. Zool. 301, 271–279 (2017).
3. Bennet-Clark, H. C. & Bailey, W. J. Ticking of the clockwork cricket: the role of the escapement mechanism. J Exp Biol 205, 613–625 (2002).
この記事を書いた人

-
ダルハウジー大学理学部心理学・神経科学科博士研究員。2017年3月東京大学大学院薬学系研究科薬学博士過程修了、博士(薬学)。2017年4月より現職。日本学術振興会海外特別研究員。2016年薬剤師免許取得。
これまで主に、カイコを用いた病原性大腸菌O-157による病原性遺伝子の機能解析および熱処理菌体による感染抵抗性獲得メカニズムの研究、並びにコオロギを用いた求愛行動の解析を行ってきました。現在は、2000年に発見された新種のコオロギを用いて、免疫経路と生殖行動の機能的な連関について探っています。また、ごく最近ではコンピュータによる画像認識の技術を応用し、コオロギの翅の模様を自動で数値化するアルゴリズムの開発を行っています。