昆虫類の体壁、そして翅関節の一部は肢の付け根に由来する
昆虫類の陸上進出を支えた「側板」
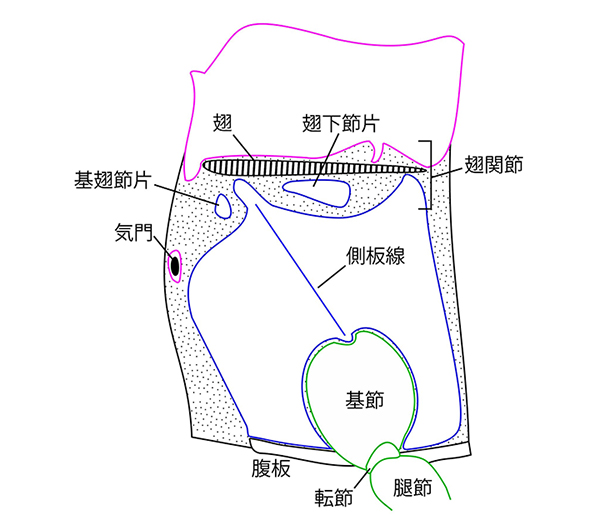
昆虫類の体は、頭・胸・腹という3つのユニットで構成され、それぞれが摂食・統合、移動、消化・生殖という機能に特化しています。翅や肢を備え、移動に特化した胸部は、背板、腹板、側板と呼ばれる3つの硬化部で構成されています。
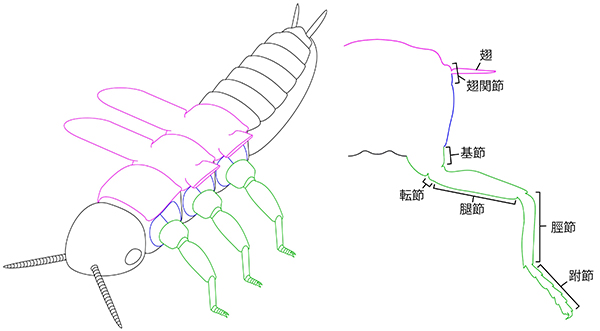
昆虫類は、長い間多足類(ムカデやヤスデの仲間)と近縁だと考えられてきましたが、現在では水生の甲殻類の一部から進化してきたことが明らかとなっています。水中から陸上へ生活の場を移す際にはさまざまな環境の変化に晒されますが、そのなかでも大きな違いのひとつが浮力の有無です。側板は、近縁な甲殻類にはみられない昆虫類の派生形質であり、水の浮力から離れ陸上に進出した昆虫類の体を支える頑健な体壁として、また歩行のための筋肉の付着点として、大きな役割を果たしています。では、この側板は、いったい何に由来しているのでしょうか。
亜基節由来説の検証 – 2μmの薄くて分厚い障壁
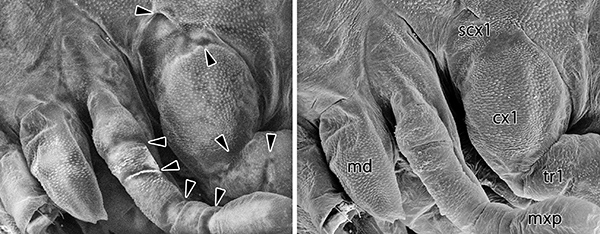
亜基節由来説、つまり側板が、付属肢の根元にあたる基節のさらに基部側にある肢環節“亜基節”に由来していることが初めて提唱されたのは、19世紀末のことです。しかしその後は、走査型電子顕微鏡(以下、SEM)を用いたいくつかの先行研究から発生学的証拠が提示されはしたものの、実はそれぞれの研究で背板・亜基節と同定されている部位が異なるという問題を抱えていました。我々は、こうした部位同定の問題が胚発生終盤の形態観察の難しさに起因していることに気が付きました。胚発生終盤になると胚は、表皮から分泌された胚クチクラと呼ばれる薄膜に包まれますが、この胚クチクラがSEMを用いた観察の大きな障害となっていたのです。
SEMというのは、高真空下に置いた試料に電子線を当て、そこから放射される二次電子・反射電子を検出することで、対象の表面構造を観察することができるという代物です。一般的なSEM観察では、前処理として試料を完全に乾燥させ、電子線による試料の帯電を防ぐために金属コーティングなどを施します。胚クチクラという柔らかい薄膜に包まれた状態の胚を乾燥させると、試料の収縮は避けられません。そこに金属コーティングを施せば、観察できるのは皺だらけになった胚クチクラの表面構造です。先行研究では、この胚発生終盤という重要なステージの形態形成を正確に追跡することができずにいたために、解釈の混乱が生じていたというわけです。
SEMで胚クチクラを「透かして」観る
金属コーティングは、胚クチクラに包まれた胚の正確な観察を妨げてしまいます。かといって金属コーティングを施さなければ、試料は瞬く間に帯電してしまい、観察どころではなくなります。この問題を解決したのが、低真空SEMを利用した観察法でした。原理の詳細は省きますが、低真空SEMの大きな利点のひとつが、帯電が起こりにくいため金属コーティングを施さずに試料を観察できる点です(デメリットとして解像度がやや低下します)。さらに、クチクラのような薄膜に、コーティングなしの状態で強い電子線を当てると、表面構造だけでなく、薄膜の直下の構造まで透過して観察することができるのです。SEMといえば、一般的には表面構造を観察するためのものですが、この手法を利用することで、胚クチクラの下で進む形態形成の詳細を追跡することが可能となり、胚発生全体をこれまでよりも高い解像度で観察することができるようになりました。また、胚の完成形である1齢幼虫においても、クチクラが柔らかいために前処理による収縮が観察の障害となっていましたが、この問題も近年開発されたナノスーツ法を導入したことで、試料を乾燥させる必要がなくなり、より正確な観察が可能となりました。
亜基節由来説の立証、そして昆虫の翅の起源へ
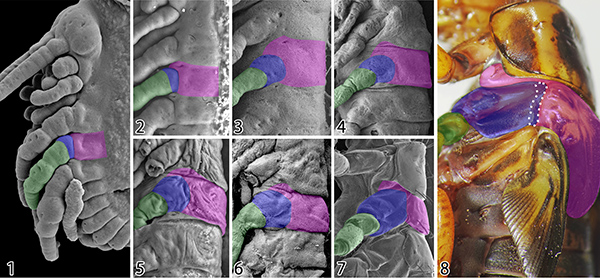
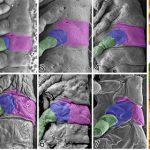
観察時の障害を乗り越えてしまえば、あとは各発生段階の形態形成過程を丹念に観察するのみです。ランドマークとなる構造を新たに見つけ、それらを頼りに形態形成過程を追跡した結果、背板ー肢境界を確定することができ、側板領域は付属肢の最基節である亜基節に由来していることがわかりました。そして、議論を重ねた結果、この成果は長年続いてきた側板形成の問題にとどまらず、翅の起源に関わるものであるということに気が付きました。
昆虫類の進化史のなかでも翅の起源は特に重要視され、長い間議論が続いてきましたが、主要な説としては「側背板起源説」と「肢(または鰓)起源説」の2つに大別されます。前者は、翅が背板の側縁部(=側背板)に由来するというもので、翅のシート状の形状・位置・気管と翅脈の類似性などから支持される一方、筋肉などの由来が説明できないという欠点を抱えていました。これに対し後者は、翅が付属肢の外側にみられる可動性の付属物(カゲロウ目幼虫の鰓など)に由来するというものですが、可動性の構造という類似性はあるものの、現存する翅とは位置や形状に大きく隔たりがあることが説明できていませんでした。また、どちらかの説を支持するような中間形の化石記録がないことも大きな問題となっていました。
そうしたなかで、近年の分子発生学的研究から提唱されたのが、両仮説を統合した「二元起源説」です。これは、側背板・付属肢外側の付属物の双方に関わる遺伝子が共同で働いたことで、翅が急速に進化したというものです。形態学からも、側板上方の一部が分離した構造が翅関節の一部を構成していることが示唆されたことはありましたが、これまでは側板の由来が定かではなかったためにそれ以上の議論に発展することはありませんでした。しかし、今回の我々の研究が、側板は付属肢の最基節である亜基節に由来していることを明らかにしたことで、翅関節の構成要素には付属肢要素が含まれており、「二元起源説」が初めて形態学的にも支持されることになりました。
今後の展開
本研究では、材料として扱いやすく、比較的派生的な系統であるフタホシコオロギを材料として用いましたが、原始的な有翅昆虫類や無翅昆虫類においても同様の側板の形成様式がみられるのか確かめる必要があります。また、陸上進出は節足動物の複数の系統で独立に起きたとされています。多足類などにもみられる側板などについても今回のようなアプローチで調べることで、新たな発見があるかもしれません。
参考文献
Mashimo, Y., Machida, R. (2017), Embryological evidence substantiates the subcoxal theory on the origin of pleuron in insects. Scientific Reports, 7, 12597. doi: 10.1038/s41598-017-12728-2
Clark-Hachtel, C.M., Tomoyasu, Y. (2016), Exploring the origin of insect wings from an evo-devo perspective. Current Opinion in Insect Science, 13, 77–85. doi: 10.1016/j.cois.2015.12.005
Takaku, Y., Suzuki, H., Ohta, I., Ishii, D., Muranaka, Y., Shimomura, M., Hariyama, T. (2013), A thin polymer membrane, nano-suit, enhancing survival across the continuum between air and high vacuum. Proceedings of the National Academy of Sciences, 110(19), 7631–7635. doi: 10.1073/pnas.1221341110
この記事を書いた人

- 福島大学大学院 共生システム理工学研究科 日本学術振興会特別研究員PD。博士(理学)。1987年群馬生まれ。筑波大学大学院 生命環境科学研究科 生物科学専攻 博士後期課程修了後、筑波大学山岳科学センター菅平高原実験所(前 菅平高原実験センター)研究員・技術補佐員を経て現職。専門は昆虫形態学・比較発生学。ジュズヒゲムシの系統・分類・発生学的研究をしつつ、現在は昆虫類の卵膜と受精様式の進化の研究をしている。
この投稿者の最近の記事
 研究成果2017年10月19日昆虫類の体壁、そして翅関節の一部は肢の付け根に由来する
研究成果2017年10月19日昆虫類の体壁、そして翅関節の一部は肢の付け根に由来する


