哺乳類の細胞分裂を守る染色体領域でRNAが果たす役割とは?
細胞分裂を守る染色体領域:ペリセントロメア
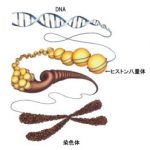
ひとつの細胞が持っているゲノム情報は、マウスで約25億塩基対、ヒトで約29億塩基対からなり、およそ2メートルくらいの長大な二重らせん構造をとっています。この長大なDNAが絡み合うことなく核内に機能的に収納できるようにするため、ヒストンH2A、H2B、H3、H4が2分子ずつ集まったヒストン8量体にDNA約146bpが巻きつき、これらがコンパクトに折りたたまれることで染色体(クロマチン)を構成しています。
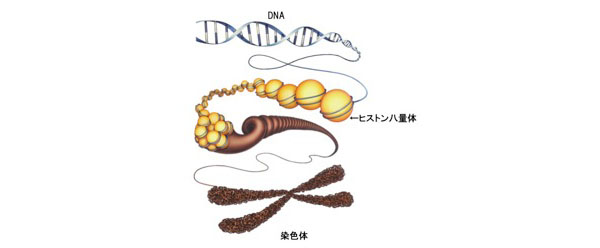
染色体にはDNAの密度が低い、つまりDNAが緩んでいて、転写が活性なユークロマチンと、DNA密度が高くDNAが凝縮することで、転写が抑えられているヘテロクロマチンに大別されます。高度に凝縮されたヘテロクロマチンは、染色体機能の維持、発生や疾患におけるエピジェネティックな遺伝子発現抑制など、さまざまな生命現象に重要な役割を果たす高次クロマチン構造です。
哺乳類の染色体では、メジャーサテライトと呼ばれる配列が1万回近くも繰り返して並んだ配列から構成されるペリセントロメア領域が動原体(セントロメア)近傍に存在し、その領域に典型的なヘテロクロマチン(ペリセントロメアヘテロクロマチン)が形成されます。まったく遺伝子の存在しないペリセントロメアは必要なのでしょうか? 実はこのペリセントロメアヘテロクロマチンは染色体が分離する際に染色体同士を結合しているコヒーシンと呼ばれる分子が最後まで残って結合している場所であり、無事に細胞分裂が起こるように守っている領域なのです。
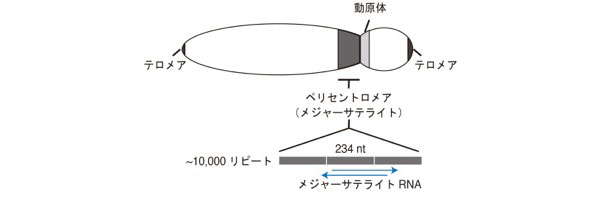
哺乳類のペリセントロメアヘテロクロマチン形成機構
哺乳類のヘテロクロマチン領域では、翻訳後修飾のひとつであるヒストンH3の9番目のリジン残基がトリメチル化されたH3K9me3が特徴的に蓄積しています。ペリセントロメアヘテロクロマチンは、ヒストンメチル化酵素のSuv39h1がこのH3K9のトリメチル化を触媒し、そのメチル化修飾部位をヘテロクロマチン構造タンパク質であるHP1が認識・結合することにより形成されることが知られています。このヘテロクロマチンの形成や機能の維持に中心的な役割を果たすSuv39hをノックアウトしたマウスでは染色体異常が誘導され、多くは致死になることが知られています。しかし、どのようにしてSuv39h1やHP1がペリセントロメア領域に存在しているのかなど、その形成の仕組みの詳細はわかっていませんでした。
ペリセントロメアからもRNAが転写される
遺伝子からRNAが転写されてそれがタンパク質に翻訳されるというセントラルドグマはよく知られています。しかし、実際にタンパク質をコードしている遺伝子は全ゲノム中の1.5%にも満たないにも関わらず、ゲノムから転写されているRNAは約7割にもおよぶことが明らかになり、このようなRNAをノンコーディングRNAと呼ぶようになりました。先ほど説明したペリセントロメア領域からもわずかながらメジャーサテライトRNAというノンコーディングRNAが転写されます。ショウジョウバエや分裂酵母などのモデル生物では、この転写されたRNAを介したRNA干渉がヘテロクロマチン形成に関与していることが知られていましたが、哺乳類では、RNAがヘテロクロマチン形成や維持に関与しているかはわかっていませんでした。
ヘテロクロマチン構造の形成に関わる因子は進化的によく保存されており、分裂酵母には、Clr4と呼ばれるマウスのヒストンメチル化酵素Suv39h1の相同因子があります。私たちは以前の研究から、Clr4がメチル化ヒストンの結合に関わるクロモドメイン(CD)を介してRNAと結合できることを見いだしていました。
そこで、Suv39h1もCDを介してRNAに結合できるのではないかと考え、生化学的に実験を行いました。その結果、Suv39h1のCDはRNAと結合し、ヒストンのメチル化の程度に影響されないことを発見しました。さらに、マウスの細胞内においても、Suv39h1はCDのRNA結合活性依存的に、ペリセントロメア領域から転写された「メジャーサテライトRNA」と結合していることがわかりました。
RNAと結合できないとヘテロクロマチン形成が遅れる
Suv39h1のCDはRNAと結合できることがわかりましたが、CDはH3K9me3とも結合できます。そこで、H3K9me3と結合するSuv39h1のCDを解析し、その結合に関わるアミノ酸残基を同定しました。そして、生化学的な解析により、両者の結合は独立していることを見いだしました。これらの結果は、Suv39h1がCDを介して、H3K9me3とRNAの両方に結合でき、両者の結合はそれぞれ独立に機能していることを示しています。では、実際に細胞内ではSuv39h1のRNA結合がヘテロクロマチン形成にどのような役割を果たしているのでしょうか?
その答えを導くために、私たちはペリセントロメア領域のH3K9me3が消失しているSuv39h欠損線維芽細胞に、変異型のSuv39h1を導入して機能解析を行いました。その結果、H3K9me3結合部位に変異を導入した細胞だけでなく、RNA結合部位に変異を導入した細胞でもヘテロクロマチン形成が遅れることがわかりました。また、これらの細胞株における核の状態を顕微鏡で観察したところ、Suv39h1のH3K9me3結合部位とRNA結合部位に変異を導入した細胞では、ヘテロクロマチン領域にSuv39h1が蓄積している細胞の割合が低下していました。
ペリセントロメアからのRNAの役割は?
これらの結果は、変異型のSuv39h1が安定的にヘテロクロマチン領域につなぎ止められていないからだと考え、Suv39h1の動的な挙動を顕微鏡で観察しました。すると、H3K9me3結合部位に変異を導入した細胞だけでなく、RNA結合部位に変異を導入した細胞も、ヘテロクロマチン領域でのSuv39h1の動きが速くなっていること、つまり可動性が増していることがわかりました。
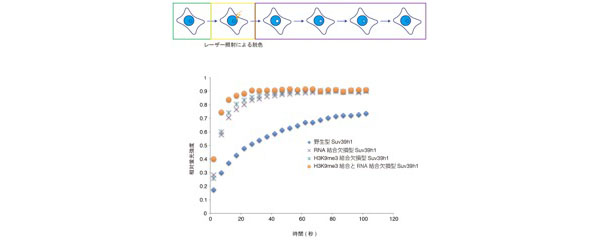
しかし、これだけではメジャーサテライトから転写されているRNAがSuv39h1の動きに影響を与えている直接的な結果にはなりません。そこで、核内のメジャーサテライトRNAをノックダウンした細胞で、Suv39h1の挙動を顕微鏡で観察したところ、ノックダウンすることでもSuv39h1の動きが速くなることを明らかにしました。
以上の結果より、Suv39h1が安定的に染色体に結合するためにメジャーサテライトRNAが重要な役割を果たし、それがヘテロクロマチン形成に重要であるということを明らかにしました。
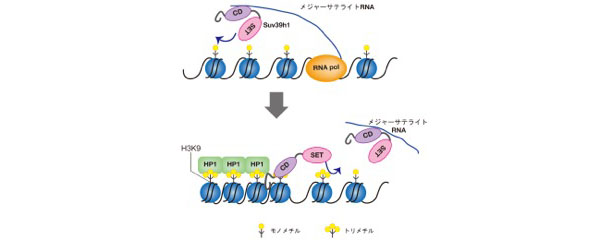
今回私たちは、哺乳類では、クロモドメイン(CD)とSETドメイン(SET)を持つSuv39h1が、ノンコーディングRNAとCDを介して結合し、またH3K9me3と結合することで、効率的にヒストンH3K9をトリメチル化し、ヘテロクロマチンを形成するという新しい機構を示しました。
近年、染色体異常が原因の先天性疾患が報告されていますが、その仕組みを完全に理解するには至っていません。進化的にノンコーディングRNAを介したヘテロクロマチン形成が保存されているという事実は、今後、未だに明らかにされていない初期のヘテロクロマチン形成を解明する糸口となり、先天性異常を引き起こす染色体不安定性を解明する重要な一歩になると期待しています。
参考文献
Atsuko Shirai, Takayuki Kawaguchi, Hideaki Shimojo, Daisuke Muramatsu, Mayumi, Ishida-Yonetani, Yoshifumi Nishimura, Hiroshi Kimura, Jun-ichi Nakayama and Yoichi Shinkai, “Impact of nucleic acid and methylated H3K9 binding activities of Suv39h1 on its heterochromatin assembly”, eLife, doi:10.7554/eLife.25317
Ishida et.al. Molecular Cell, 47, 228-241 (2012)
この記事を書いた人

-
眞貝洋一(写真左)
1990年順天堂大学大学院医学研究科博士課程修了、博士(医学)。コロンビア大、ハーバード大博士研究員、日本ロシュ研究所主任研究員、京都大学ウイルス研究所准教授、教授を経て、2011年より現職。2001年、当時その機能がよくわかっていなかったG9aという分子がヒストンのリシンメチル化酵素であることを見出し、その後は一貫してヒストンメチル化による細胞記憶メカニズムの研究を進めている。
白井温子(写真右)
2007年東京大学大学院農学生命科学研究科博士課程終了、博士(農学)。理化学研究所 協力研究員、理化学研究所 発生・再生科学総合研究センター 研究員、基礎科学特別研究員を経て、2012年より現職。高次クロマチンが形成される機構を解明すべく研究を行っています。
この投稿者の最近の記事
 研究成果2017年9月6日哺乳類の細胞分裂を守る染色体領域でRNAが果たす役割とは?
研究成果2017年9月6日哺乳類の細胞分裂を守る染色体領域でRNAが果たす役割とは?


