ヘビの胴体が長いメカニズムがわかった! – 脊椎動物の後ろ足の位置の多様性を生み出す仕組み
私たちの後ろ足は背骨の中の仙椎の場所にあります
私たちヒトを含む脊椎動物の体の中心には背骨(せぼね)があります。背骨はたくさんの脊椎骨(せきついこつ)が1列に並んだ構造をしており、脊椎骨は形の違いで頭に近い方から頸椎(けいつい)、胸椎(きょうつい)、腰椎(ようつい)、仙椎(せんつい)、尾椎(びつい)と呼ばれています。私たちの後ろ足は骨盤(こつばん)を介して仙椎に接続しています。
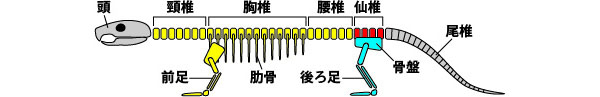
後ろ足(水色)は仙椎(赤色)の場所に作られている
仙椎と後肢の位置関係は進化の過程で保存されています
さまざまな動物の骨格を見てみると、今生きている動物だけでなくすでに絶滅してしまった恐竜や首長竜、ヘビの祖先で手足を持つテトラポドピスに至るまで、あらゆる生物種において後ろ足は仙椎に接続していることがわかります。このように、仙椎の場所に後ろ足が形成されるメカニズムは進化の過程で非常に良く保存されています。興味深いことに、進化の過程で脊椎骨の数は大きく変化していますが、後ろ足は必ず仙椎の位置に作られます。
これまで、なぜ後ろ足は必ず仙椎の場所に作られるのか、また進化の過程でどのようにして後ろ足の位置が多様化していったのかはまったくわかっていませんでした。
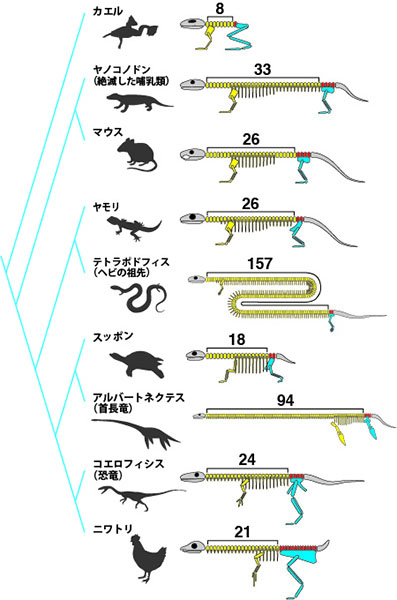
進化の過程を見ても後ろ足(水色)は必ず仙椎(赤色)の場所に作られている
GDF11が作用した場所に仙椎と後ろ足が作られる
“進化”は、受精卵から体がつくられ産まれる直前までの状態である「胚」の“発生過程”の変化の蓄積によって起こります。私たちは、さまざまな脊椎動物において後ろ足ができるときの発生過程を調べれば、なぜ後ろ足が必ず仙椎の場所に作られるのか、また進化の過程でどのようにして後ろ足の位置が多様化していったのかを、明らかにできるのではないかと考えました。
まず私たちは、体の発生過程を観察しやすいニワトリの胚を用いて後ろ足の発生メカニズムを詳細に調べました。その結果、背骨が作られる場所の中でGDF11(ジーディーエフイレブン)と呼ばれるタンパク質が働き始めた場所が、将来の仙椎になることがわかりました。さらにGDF11タンパク質は仙椎になる組織の隣の組織(専門用語で側板中胚葉という名前の組織です)にも働きかけて、そこに後ろ足と骨盤をつくることを発見しました。この発見により、脊椎動物の後ろ足が必ず仙椎の位置に作られているメカニズムが初めて明らかにされました。
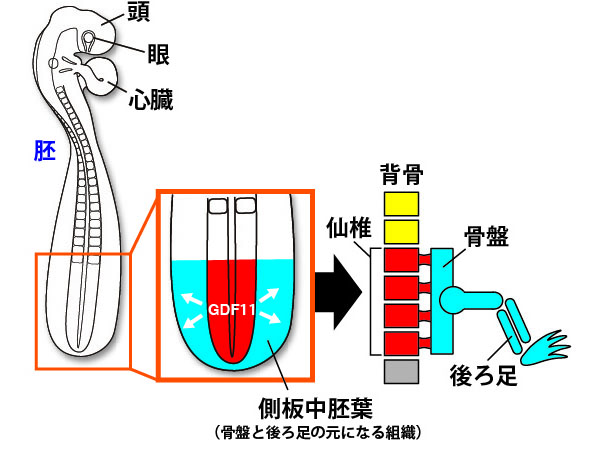
背骨が作られる場所の中でGDF11と呼ばれるタンパク質が働き始めた場所が、将来の仙椎になり、隣接する組織に後ろ足が形成される
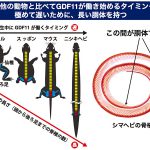
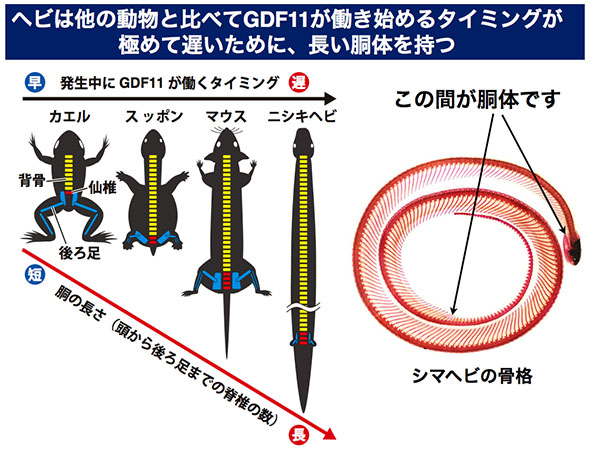
ヘビはGDF11が働き始めるタイミングが極めて遅いために長い胴体を持つ
次に私たちは、動物種間で後ろ足の位置の違いが生まれる仕組みを調べるために、脊椎動物の中で胴体が短い(頭から後ろ足までが近い)ものと、胴体が長い(頭から後ろ足までが遠い)もの、合わせて9種の動物においてGDF11の働き方を調べました。
その結果、カエルやカメなどの胴体が短い(頭から後ろ足までが近い)ものは発生中にGDF11が働き始めるタイミングが早く、エミュー(鳥の仲間)やヘビなどの頭から後ろ足までが遠いものではGDF11が働き始めるタイミングが遅いことがわかりました。
この結果から、進化の過程で後ろ足の位置が多様化していった原因は、GDF11というたったひとつの遺伝子から作られるタンパク質の発生中に働くタイミングが異なるためであることが明らかになりました。特にヘビは、他の動物と比べてGDF11が働き始めるタイミングが極めて遅いために、長い胴体を持つことがわかりました。
今後の期待
GDF11は、ヒトを含むすべての脊椎動物が持っています。よって、地球上に存在する多様な形態を持つ脊椎動物すべてにおいて、後ろ足の位置の多様性はGDF11というたったひとつの遺伝子から作られるタンパク質が働くタイミングが胎児期に異なることで生み出されたと考えられます。特にヘビは、他の動物と比べてGDF11が働き始めるタイミングが極めて遅いために、長い胴体を持つことがわかりました。
後ろ足の位置の多様性に代表される生物の大進化は、これまで進化学の分野では体の形を作るHox遺伝子(ホックス遺伝子)の変化によって引き起こされたと考えられて来ました。今回の研究で、GDF11はHox遺伝子の働く場所をまさに制御している働きを持つことがわかりました。これにより生物の形態の大進化は、オーケストラの指揮者の様に働く、思った以上に少数の遺伝子の変化によってもたらされた可能性が推測されます。
GDF11の機能を阻害すると胴が長くなり、体の下半身全体の位置がずれることがわかっています。今後は胎児期にGDF11の作用するメカニズムをさらに調べていくことで、仙椎や後ろ足だけではなく、下半身全体の器官の位置を決める発生メカニズムの解明に大きく貢献することが期待されます。
参考文献
Matsubara Y., Hirasawa T., Egawa S., Hattori A., Suganuma T., Kohara Y., Nagai T., Tamura K., Kuratani S., Kuroiwa A., Suzuki T. (2017), Anatomical integration of the sacral-hindlimb unit coordinated by GDF11 underlies variation in hindlimb positioning in tetrapods. Nature Ecology and Evolution, 1, DOI: 10.1038/s41559-017-0247-y
この記事を書いた人

- 名古屋大学大学院理学研究科生命理学専攻 講師。静岡県出身。慶應義塾大学理工学部応用化学科卒業、奈良先端科学技術大学院大学バイオサイエンス研究科後期博士課程修了 博士(バイオサイエンス)。日本学術振興会特別研究員、海外特別研究員(於ウィスコンシン大学)、さきがけ研究員(兼任)などを経て2010年より名古屋大学大学院理学研究科生命理学専攻(助教)、16年より同(講師)