酢酸を使って乾燥ストレスに打ち勝つ植物の生存戦略とは
環境応答とエピジェネティック制御
あらゆる生命活動の基本となる「水」の欠乏は、生物にとってクリティカルヒットになります。地球温暖化にともなう気候変動により、世界各地で長・短期に関わらず乾燥や干ばつが起こり、作物生産や緑資源の壊滅的な被害が年々拡大しています。私たちは、乾燥への対抗策を見出すため、基礎研究の立場から植物の乾燥耐性・適応機構の解明を進めています。
今回は、これまでの研究では知りえなかった、まったく新しい植物の乾燥ストレス応答機構を制御するキーレギュレーターの同定とその作用機序の解明を目指して、遺伝子の応答制御と関連の深いクロマチン修飾変動(特にヒストン修飾)に目をつけ、解析を行いました。ヒストン修飾は真核生物に保存されるエピジェネティック情報のひとつです。モデル植物であるシロイヌナズナを用いて、ヒストン修飾酵素をコードする遺伝子の変異株を多数集め、乾燥ストレス下での耐性・感受性を調べるスクリーニングを行いました。その結果、ヒストン脱アセチル化酵素HDA6の遺伝子変異植物体が、著しく強い乾燥耐性を示すことが明らかとなりました。

hda6遺伝子変異株および野生型親株のシロイヌナズナに対し、給水停止により2週間乾燥処理したのち、3日間の再給水を行ったときの生存を確認した
乾燥によるダイナミックな代謝変換
hda6遺伝子変異体とその親株の野生型植物を用いて、乾燥条件下でのゲノムワイドな遺伝子発現変動、およびこれにリンクした代謝物の変化を調べました。その結果、hda6遺伝子変異体と野生株ではともに、「乾燥処理により中心代謝系である解糖系が大々的に抑制」される一方で、解糖系の中間代謝物であるピルビン酸を出発点とする代謝バイパス経路の「酢酸合成系が乾燥により誘導・活性化」されることが明らかとなりました。次に乾燥条件下での植物体内の酢酸蓄積量を調べたところ、hda6遺伝子変異体では野生型のシロイヌナズナに比べ、およそ2倍量の酢酸が蓄積していました。
さらに生化学的解析の結果から、HDA6タンパク質は酢酸合成に必須の酵素をコードする2つの遺伝子領域に結合し、これら遺伝子の転写活性を直接制御していることがわかりました。つまり、乾燥に応答して植物体内では解糖系から酢酸合成系へのダイナミックな代謝変換が起こり、その変換スイッチ分子として植物のエピジェネティック因子であるHDA6が働いていることが明らかとなりました。
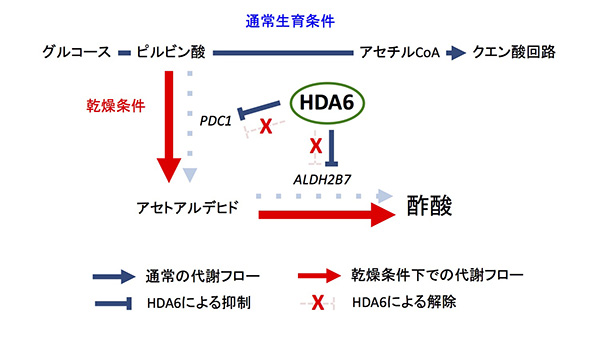
植物において、通常生育時に解糖系からクエン酸回路への代謝フローが機能している。このとき、HDA6タンパク質は酢酸合成に働く酵素遺伝子PDC1とALDH2B7を直接抑制している。乾燥条件下では、解糖系からクエン酸回路へのフローが抑制されるとともに、これら遺伝子領域からHDA6タンパク質が乖離し、酢酸合成系の抑制が解除されることで、乾燥に応答した酢酸の合成が誘導促進される。
酢酸が植物に乾燥耐性を付与するメカニズム
では、どのようにして酢酸は植物に乾燥耐性を与えることができるのでしょうか? まず私たちは、植物体の外部から酢酸を投与することで、植物に乾燥耐性を付与できるかどうか調べました。酢酸とその他の酸溶液(塩酸、蟻酸、酪酸、乳酸、クエン酸)を用いてシロイヌナズナに9日間前処理し、その後、およそ2週間給水を停止して植物を乾燥状態においたときの生存について調べました。その結果、今回用いた酸溶液の中では酢酸だけがシロイヌナズナに強い乾燥耐性を付与でき、乾燥環境下での植物の生存にポジティブに働くことがわかりました。
次に酢酸の作用機序を明らかにするため、外部からシロイヌナズナに酢酸を与えたときの植物ホルモンの量的変化などを調べた結果、酢酸投与により植物体内のジャスモン酸量が一過的に急上昇することがわかりました。ジャスモン酸は植物の傷害応答に関与することが知られている植物ホルモンです。
さらに、酢酸と乾燥の複合的な処理により、ジャスモン酸によって誘導を受ける遺伝子群が高感度に発現誘導されること、および、ジャスモン酸合成とシグナル伝達に関わるタンパク質をコードする遺伝子の破壊株では、乾燥に著しく弱くなることなどもわかりました。
これらにより、「酢酸がジャスモン酸合成を誘導し、その結果、傷害応答に関わるジャスモン酸シグナル伝達系の下流遺伝子群が活性化されることで、植物は乾燥に強くなる」というメカニズムが明らかになりました。

2週齢のシロイヌナズナ野生株に10mMのさまざまな酸溶液を用いて前処理をした後、およそ2週間の乾燥処理を施した。乾燥処理後、3日間の再給水を行い、植物の生存の様子を調べた。10mM酢酸で処理したシロイヌナズナの野生型植物体だけが顕著な乾燥耐性を示した。これにより、酢酸による植物への乾燥耐性付与効果が確認できた。
酢酸の役割はもうひとつある
生体内の酢酸はアセチルCoAに代謝変換されることが知られています。このアセチルCoAはクロマチンの活性化に必要なヒストンアセチル化修飾の唯一の基質と考えられています。私たちは、酢酸投与によるゲノム活性化の可能性を調べるため、放射性同位体でラベルした酢酸を植物体内に取り込ませた後、ヒストンタンパク質を精製して、ラベルした酢酸がヒストンタンパク質へアセチル化修飾として取り込まれているか否かなどを調べました。
その結果、外部から与えた酢酸は、生体内のヒストンH4に取り込まれるだけでなく、ゲノムワイドなヒストンアセチル化を促進してクロマチン・遺伝子群の活性化を促進すること、また、それら遺伝子群のなかには酢酸によって誘導を受けるジャスモン酸下流の遺伝子が多数含まれることがわかりました。
このことから、今回見つかった植物の新規乾燥耐性獲得メカニズムにおいて、酢酸はジャスモン酸の一過的合成と下流遺伝子発現の誘導、およびこれら遺伝子群に対するクロマチンレベルでの活性化という二重の活性化経路で複合的に機能していることが明らかになりました。
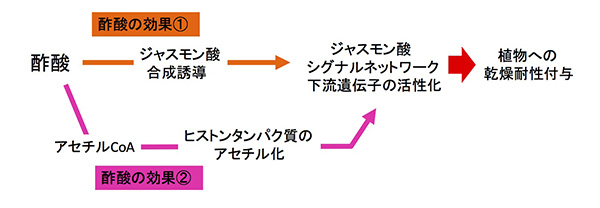
酢酸の効果は二重の経路で植物に乾燥耐性を付与する。酢酸の効果①:酢酸の投与により、傷害応答に働く植物ホルモンであるジャスモン酸の合成が一過的に誘導される。合成されたジャスモン酸の刺激により、ジャスモン酸シグナルネットワーク下流の遺伝子群が活性化される。酢酸の効果②:同時に、投与された酢酸は植物体内でアセチル基としてヒストンタンパク質に取り込まれ、①で活性化されるジャスモン酸シグナルネットワーク下流標的遺伝子の活性化誘導に機能する。
進化的に植物に保存されたメカニズム
私たちは、乾燥条件下で酢酸合成を担う2つの遺伝子(PDC1とALDH2B7)を同定しています。進化的系統解析の結果から、ピルビン酸からアセトアルデヒドへの反応を触媒する酵素遺伝子PDC1は酵母および植物に特異的に保存されており、一方で、アセトアルデヒドから酢酸への触媒酵素遺伝子ALDH2B7は酵母からヒトに至るまで高度に保存されていました。このことから、乾燥耐性に関わる酢酸合成系の進化的な保存性は、植物に特化したものであると推察されました。
この推察をもとに、シロイヌナズナ以外の植物種において酢酸による乾燥耐性付与効果を明らかにするため、イネ、コムギ、トウモロコシおよびナタネを用いて、乾燥実験を行ったところ、単子葉植物、双子葉植物を問わず実験に用いたこれら全ての植物種で、酢酸を与えることにより乾燥に強くすることができました。

トウモロコシ、イネ、ナタネ、コムギに、0-50mM酢酸で前処理を施したときの、乾燥耐性付与効果を調べた。単子葉、双子葉に関わらず、酢酸処理による乾燥耐性の付与効果が確認できた。また、すべての植物種において、有効な酢酸濃度はおよそ10-30mMの範囲であることが示された。
今後の展開
今回、私たちが発見した「エピジェネティック因子により制御される、酢酸—ジャスモン酸を介した植物の乾燥耐性機構」は、これまでに報告されている多くの乾燥応答遺伝子との関連性がほとんどなく、植物の乾燥応答機構と環境変動時の生存戦略を考えるうえでも、まったくの新しい知見と可能性を拓くものだと言えます。
今後は、ジャスモン酸により活性化される下流遺伝子ネットワークの解析を通して、植物に乾燥耐性を与えることができるジャスモン酸ネットワーク下流の実行因子(遺伝子や生体分子など)の解明を進めたいと考えています。また現段階において本研究成果は、酢酸による植物への乾燥耐性付与効果を「実験室内」で明らかにできたというだけであり、農地の乾燥対策や砂漠の緑化にすぐに適用できるものではありません。農地での応用を見据えた研究・開発および実証実験を進めていくことで、干ばつ、乾燥による地球規模での植物資源の減少を食い止めるための技術の確立を、私たちは目指しています。
参考文献
Kim JM, To TK, et al. Nature Plants 2017, article number: 17097. doi: 10.1038/nplants.2017.97.
この記事を書いた人

-
金 鍾明(写真中央)
理化学研究所 環境資源科学研究センター 研究員。長崎大学水産学部卒業、奈良先端大学院大学バイオサイエンス研究科博士後期課程修了、アメリカ・UCLA 分子生物学研究所にてポスドク、理化学研究所 植物科学研究センター 研究員を経て、2013年より現職。酢酸による植物の乾燥耐性機構を発見。理化学研究所に入所以来、植物の環境応答とゲノム・クロマチン動態変化に関する研究を行っています。
関 原明(写真左)
理化学研究所 環境資源科学研究センター 植物ゲノム発現研究チーム チームリーダー。京都大学農学部卒業、広島大学大学院理学研究科にて博士号(理学)を取得。理化学研究所 植物分子生物学研究室 研究員などを経て、2013年より現職。植物の環境ストレス適応・耐性強化とキャッサバの分子育種に関する研究を行っています。
藤 泰子(写真右)
東京大学大学院理学系研究科 助教。東京大学理学部 生物化学科卒業、東京大学理学系研究科 修士課程修了、アメリカ合衆国UCLA 留学、東京大学理学系研究科にて博士(理学)を取得。国立遺伝学研究所特別研究員、日本学術振興会特別研究員を経て、2015年より現職。エピジェネティクス、ゲノミクス、植物遺伝学に関する研究を行っています。
この投稿者の最近の記事
 研究成果2017年8月1日酢酸を使って乾燥ストレスに打ち勝つ植物の生存戦略とは
研究成果2017年8月1日酢酸を使って乾燥ストレスに打ち勝つ植物の生存戦略とは


