ゲル成分でできた繭を作るカイコ – モンシロチョウが持つタンパク質をカイコの絹糸腺で発現
カイコとヒトの歴史
カイコ(Bombyx mori)という昆虫は、人類の歴史のなかで、人為淘汰の末にヒトが訓化してきたものであり、ヒトに寄り添って生きてきました。記録に残る時代にはすでに、カイコはヒトが飼わなければ生きられない生物となっていました。古くは、絹は交易において貨幣にも等しい重要なものとなり、数千年の歴史のなかで、絹をいかに多く生産するか、祖先が智恵をしぼってきたのです。また、日本では明治期以降の養蚕振興に伴い、多くの先輩研究者が遺伝学を用いて品種改良を行い、日本種/ヨーロッパ種/中国種の長所をあわせ持つ雑種を作り出した結果、日本ブランドと呼ぶべき高品質なシルクが生み出され、昭和初期には主要輸出品目になっていました。しかし、輸出品としてのシルクは次第にその役割を終え、現在では国内生糸産業はその黄金期に比べるとごくわずかなものとなってきています。
生糸以外で逆転の発想?
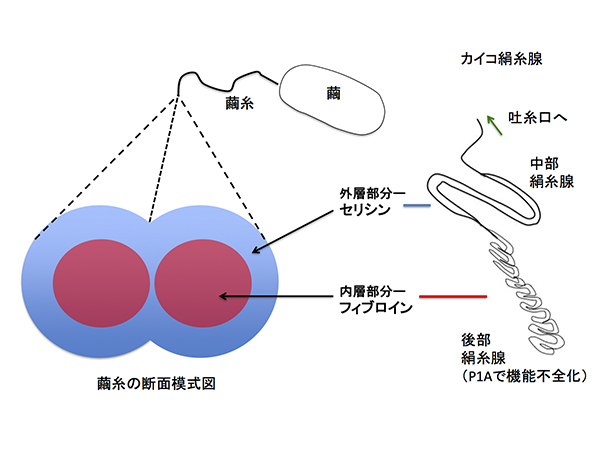
カイコの繭糸は、2層構造であり、後部絹糸腺が作るフィブロインの内層と、主として中部絹糸腺が作るセリシンの外層から構成されます。セリシンは高分子の糊状タンパク質であり、糸を繭の中でつなぎ止める役目があります。繭を高温のアルカリ性水溶液で煮ると、外層のセリシンが溶けてのぞかれ、紡ぐことで溶け残ったフィブロインの糸が取り出されます。この製糸と呼ばれる工程でセリシンの高分子構造は破壊されてしまいます。
昨今、生糸のフィブロインではなく、これまで製糸で捨てられていたセリシンにも注目が集まっています。なぜなら、細胞増殖の刺激になる性質や、化粧品材料として有効な保湿性がセリシンには期待されるからです。セリシンだけでできた繭を作るカイコを人工的な遺伝子組換えで作り出すことができないか? という発想に至り、研究を開始しました。しかし、これは、人類が取り組んできたカイコの品種改良の歴史の流れから大きく逸脱する発想でした。
モンシロチョウの細胞障害性タンパク質を利用する
フィブロインを作らないようにする遺伝子操作で、中部絹糸腺で発現するセリシンのみを吐き出すカイコを作れるのでは? と考えたものの、フィブロインの遺伝子発現を抑制する操作が簡単にできるようなゲノム編集技術がカイコではあまり進んでいません。さらに、フィブロインには2つのタンパク質サブユニットがあるので、2つ同時に遺伝子の働きをノックアウトする操作は容易ではありません。
そこで、モンシロチョウのピエリシンファミリーのタンパク質を用いることを考えました。モンシロチョウ(Pieris rapae)の蛹の血液中にピエリシン-1と呼ばれる細胞障害性タンパク質がこれまでに見つかっていました。ピエリシン-1は、ほ乳類細胞に侵入して、DNAにADP-リボースを付加することでアポトーシスを誘導するタンパク質です。ピエリシン-1の生理的な役割は十分調べられたとはいえないわけですが、おそらく、変態によって不要となる幼虫組織をのぞく役割があるのではと推察されています。
ピエリシンファミリーのタンパク質は、その1次構造が微生物の類似酵素に近く、しかもシロチョウの仲間にしか見つからないことから、もしかすると寄生した微生物のピエリシンの先祖型遺伝子をシロチョウの先祖が取り込んで利用するようになったことも考えられます。
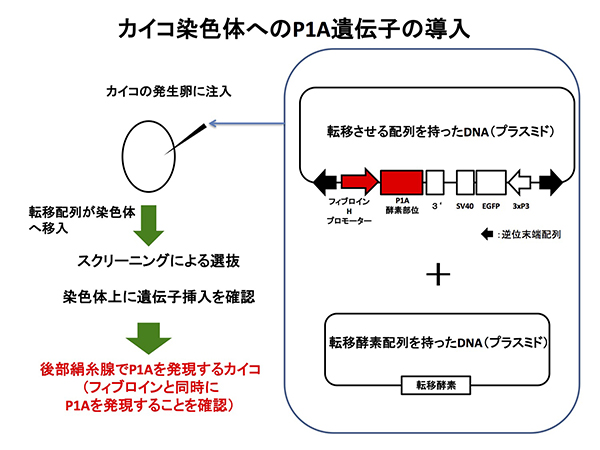
このピエリシン-1は、あまりに活性が強すぎて、これまでに生きた生物の細胞で発現に成功した例がありませんでした。これでは到底使えないわけですが、運の良いことに、ピエリシン—1A(P1A)というピエリシン—1よりも活性の強さが約5%程度のホモログがモンシロチョウから見つかりました。このP1A遺伝子に手を加えて、細胞への侵入に関わる領域をのぞいた断片をカイコの染色体へと挿入しました。
セリシン繭産生カイコ
この遺伝子組換えカイコでは、P1Aが後部絹糸腺で特異的に発現しました。また、後部絹糸腺において肥大化成長に異常をきたしたために生じる凸凹状のいびつな形態が観察されました。しかし、後部絹糸腺は細胞死による消失が見られませんでした。さらに、後部絹糸腺でフィブロインの2つのサブユニットタンパク質の発現は、転写のレベルで阻害を受けていました。
一方、普段から発現している遺伝子の転写にはP1Aはあまり影響しませんでした。絹糸腺における機能の改変のため、この遺伝子組換えカイコはほぼ100%セリシンでできた繭(セリシン繭)を作ることがわかりました。突然変異で生じたセリシン繭産生カイコがこれまでにも見つかっていましたが、これらは5〜20%程度のフィブロインを含む糸をわずかな量しか吐かないという性質を持つものであり、我々が作った遺伝子組換えカイコのようにほとんどフィブロインを含まない繭を安定的に作るカイコは今までいませんでした。

古典的な研究において、カイコの絹糸腺を幼虫期に手術により取り除くと、成虫にまで成長して子孫を作ることができなくなったという報告があります。この理由として、絹糸昆虫が幼虫期の終わりに糸を吐き出すのは、もともと幼虫期に取りすぎて余剰となった栄養分をタンパク質に変換してそれを体外へと捨てるためだと考えられてきました。絹糸腺を取り除いた場合に、本来ならば糸に組み入れて体外へと吐き出されていたアミノ酸が体内に残ってしまうことは、アミノ酸過剰の状態を招き、このため蛹期以降上手く成長できなくなるというものです。糸成分の約75%がフィブロインであるため、得られた遺伝子組換えカイコは捨てるべきアミノ酸を大量に保持する、すなわちアミノ酸過剰を伴う蛹となるはずですが、得られたカイコの全成長過程で健康上の問題は見られず、何世代もP1A遺伝子ホモの子孫を継代し続けています。このことは、手術で器官を取り除くことと、遺伝子の操作による組織の機能不全は必ずしも等しくはならないことを意味しています。
得られた遺伝子組換えカイコの有用性
得られた遺伝子組換えカイコのセリシン繭から、高濃度の塩を用いることにより高分子のセリシンが得られることがわかりました。しかも、このセリシンはゲル化させることができ、ゲルを加工することでシートやスポンジも作れることがわかりました。すでにさまざまな産業分野でセリシンは用いられているのですが、製糸工程で激しく変性しているためゲル化しません。つまり、高分子セリシンは立体的に配置しつつ水を大量に溜め込むことができるという性質のあることがわかります。この点が、さまざまな日用品や化粧品の材料として応用できないかと期待されます。
また、このセリシンは3次元のバイオマテリアルとして有用ではないかと考えられます。というのも、このセリシンのゲルの上で、ES細胞株を培養することができ、また、細胞増殖因子をゲルから放出して細胞に与える性質があることもわかりました。このように、P1Aを用いることで、既存のゲノム編集とは異なる技術で、特定組織の機能不全化を実現することができ、得られたカイコはセリシン繭をつくるという有益な形質を持つこともわかりました。このP1Aによる組織の機能不全化を他の生物でも利用できるようになれば、たとえば、花粉症対策のための花粉を作らない樹木を作り出すことも夢ではなくなるかもしれません。
参考文献
Otsuki R, Yamamoto M, Matsumoto E, Iwamoto S, Sezutsu H, Suzui M, Takaki K, Wakabayashi K, Mori H Kotani E. 2017. Bioengineered silkworms with butterfly cytotoxin-modified silk glands produce sericin cocoons with a utility for a new biomaterial. Proc. Natl. Acad. Sci. U. S. A. vol.114 (26), 6740-6745. DOI/10.1073/pnas.1703449114.
伊藤智夫.1985.カイコはなぜ繭をつくるか.講談社サイエンティフィック.
この記事を書いた人
- 京都工芸繊維大学応用生物学系に勤めております。この大学は、カイコや養蚕に関する研究・開発の伝統があり、このため、カイコの研究について存分に取り組ませていただいております。遺伝子を組換えて、新しい性質を持ったカイコ、興味ある事象を見せてくれるカイコを作ることを目標にしています。将来的には、人の生活に役立ち、産業的な価値を持つカイコを開発していきたいと思っております。



