霊長類の色覚進化の道筋を探って – 3色型色覚は顔色を見分けるのに適している?
3色型の色覚を持つ霊長類
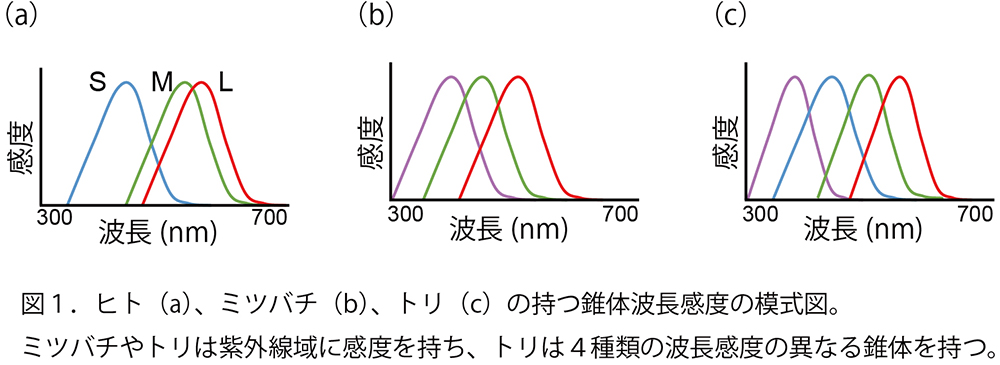
ヒトを含む多くの霊長類は、眼球の網膜に3種類の錐体細胞を持っており、それぞれ光に対する波長感度が異なります。感受性が短波長側にある方から、S(short)、M(middle)、L(long)錐体です(図1a)。ほとんどの哺乳類は、S錐体と、長波長側のもうひとつの錐体による2色型色覚です。S、M、L錐体を、青、緑、赤錐体と呼ぶこともありますが、ヒトが純粋な緑と感じる波長は、M錐体が持っている波長感度のピーク(530nm)よりも短波長側、純粋な赤と感じる波長はL錐体の波長感度のピーク(560nm)よりも長波長側にあります。これは、M、L錐体の波長感度が大きくオーバーラップしているため、一方の錐体の反応がより大きくなる波長を緑や赤と感じるためと考えられます。一般に、理想的な錐体の感度分布は、ミツバチや多くのトリにみられるような均等分布で、より多くの波長を区別できると考えられています(図1b,c)。では、霊長類のM、L錐体のオーバーラップには、何か理由があるのでしょうか?
霊長類型3色型色覚の理由とは?
あまり面白くない説明として、M、L錐体が生物進化の歴史においては比較的最近分岐したということが挙げられます。2つの錐体の違いは、有しているオプシンタンパクのわずかな違いに由来します。364個のアミノ酸からなるオプシンのたった3アミノ酸の違いで30nmの波長感度の違いが生じていることがわかっています。よって、もっと大きな波長感度の違いを起こすアミノ酸変化が起こっていないため、感度がよく似ているとも考えられます。
大きな波長感度の変化が起こらないもうひとつの説明として、光のレンズにおける屈折率が波長によって異なるために起こる、色収差と呼ばれる現象を最小にするためだという考えもあります。なぜなら、M錐体とL錐体の応答の合計が明るさ知覚に使われ、物体の輪郭など微細な空間視に関わるため、色収差が起こると輪郭がぼやけてしまうからです。このような系統的または機能的な制約のために、M、L錐体の波長感度は似通っているとも考えられます。
一方で、M、L錐体がオーバーラップしていることで、何か適応的な意義があるのかもしれません。ひとつは、長波長側の微細な波長変化を捉えられる可能性です。M、L錐体の比較により、緑、黄、橙、赤というさまざまな色の知覚を生み出すことができるのです。適応的な意義の第一の候補として、古くから、緑の葉の背景から、果実を見つけやすいことが挙げられていました(果実説)。この説を野生下のサルで検証すべく、長らく果実採食の研究を行ってきました。しかしながら、野生のサルの行動観察では、果実採食場面での3色型色覚の顕著な有効性を示すことができませんでした。むしろ、サルは明るさや嗅覚などの様々な手がかりも使って果実を採取していることがわかりました。ちょうどそのころ、顔色など、露出している肌の色の変化検出こそが3色型色覚進化の理由ではないかとする論文がMark Cangiziらによって出されました(社会的シグナル仮説)。
これらを背景として、果実説を研究していたときからの共同研究者であるAmanda Melinさん、アカゲザルの顔色変化について研究していたJames Highamさんらと今回の研究を計画しました。
顔色変化検出実験
Highamさんの研究室では、アカゲザルのメスの顔色がホルモン動態と一致して繁殖期と非繁殖期で異なることを示していました。また、オスはその変化を見分けているという行動データも示していました。さらに詳しく、霊長類型の3色型色覚が、2色型や、その他の錐体感度を持つ3色型と比較して顔色をよく見分けることができるかを、アカゲザルとほぼ同じ錐体感度分布を持っているヒトを対象として実験的に調べることにしました。Melinさんが開発していた色覚シミュレーションプログラムを使って、デジタル画像から、各色覚型が持つ色空間における色度と彩度を計算し、その色覚型で見えるであろう色の違いを一般的な3色型の人が体験できる画像に変換しました(図2)。
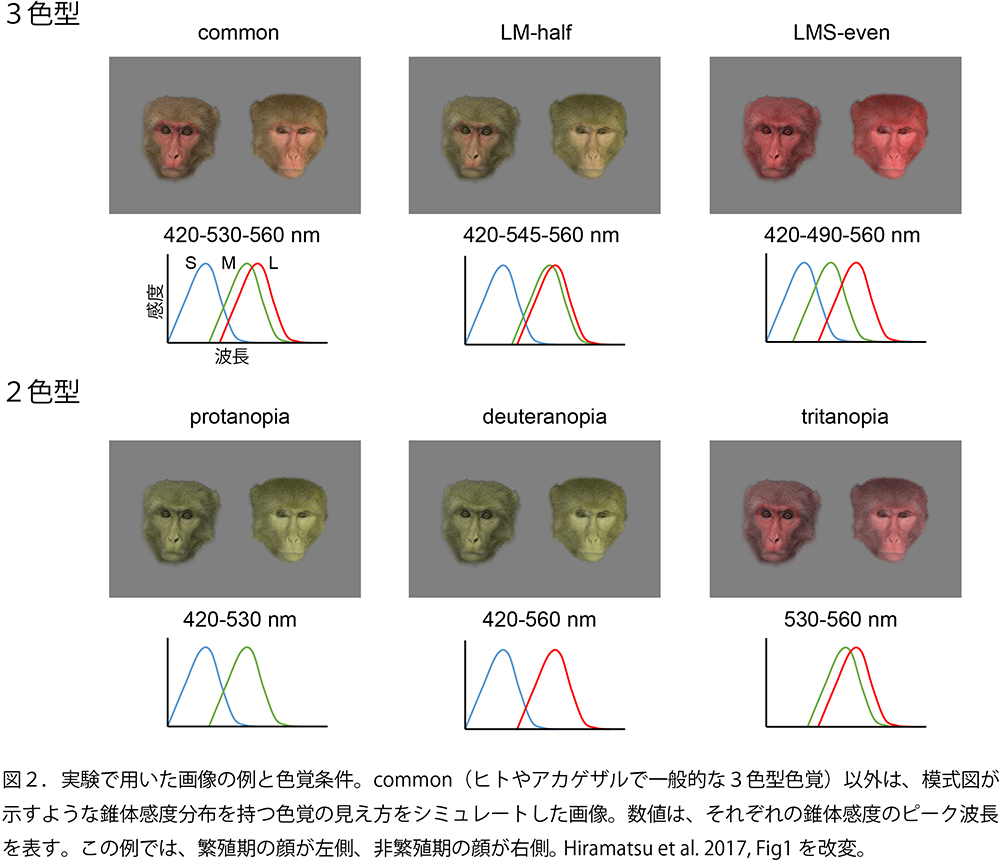
色覚型条件には、以下の6つを用いました。
1) 画像を変換していない3色型(common)
2) L錐体とM錐体のピーク波長の違いが通常の半分になった3色型(LM-half)
3) L、M、S錐体の感度が均等に分布した3色型(LMS-even)
4) L錐体を持っていない2色型(protanopia)
5) M錐体を持っていない2色型(deuteranopia)
6) S錐体を持っていない2色型(tritanopia)
3)以外は、ヒトの中にも多型として存在する色覚です。また2)は、リスザルなど、中米に生息している広鼻猿類と呼ばれるサルによくみられる3色型です。
繁殖期と非繁殖期に撮られたメスの同一個体の顔写真を左右に並べ、24頭のメスから、32枚の顔ペアを作りました。参加者にはどちらが繁殖期の顔であるかをできるだけ素早く答えてもらいました。まず、16の顔ペアを用いて、参加者の回答に応じて、正解、不正解のフィードバックを与える訓練セッションを行いました。左右のどちらが繁殖期かを変えて、合計32試行を行い、呈示する顔ペアの順序はランダムとしました。その後の5つのテストセッションでは、新しい16の顔ペアも用いました。新しい顔ペアには、正解、不正解のフィードバックを与えませんでした。各色覚条件には、それぞれ10名が参加しました。
テストセッションにおける、新しい顔ペアに対する正答率を図3に示します。
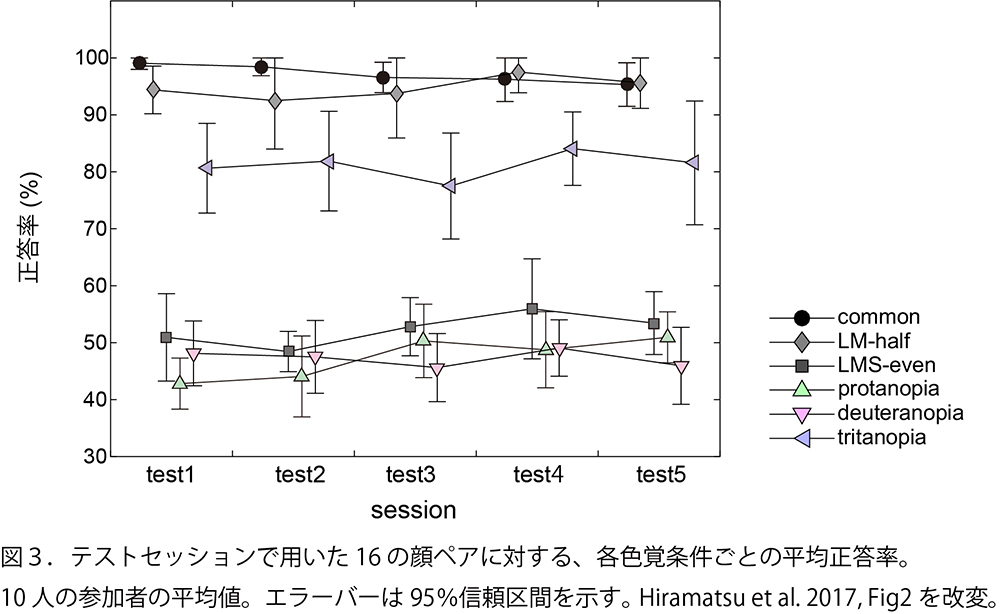
1)の通常タイプの3色型が最も正答率が高いと予測していたのですが、2)のLM-halfの3色型も1)と同様に、繁殖期と非繁殖期の顔をよく見分けることができました。3)のLMS-evenの3色型は、4)、5)の2色型と同様に、適当に答えたときの正答率である50%前後にとどまりました。長波長側にひとつの錐体しかない時には、微細な顔色の違いを検出できないためと考えられます。一方で、6)のS錐体を持っていない2色型は、80%程度の成績を示しました。
また、実験後に方略についてインタビューしたところ、正答率が高かった条件では、多くの参加者が「顔の赤さに着目した」と答えましたが、正答率が低かった条件では、目の大きさや毛の状態など、さまざまな部分に着目していたことがわかりました。このことは、色覚が異なることによって同じ画像を見ても異なる部分に着目する行動の多様性を示唆しており、現在主として進めている研究テーマにつながっています。繁殖期のサルにおいては、音声などその他の手がかりも使えるでしょうが、顔色は大きな手掛かりとなることでしょう。
今回の実験によって、霊長類が持つM、L錐体の波長感度が長波長寄りに偏ってオーバーラップしている色覚は、アカゲザルにおいて生物学的に重要と考えられる顔色変化を見分けやすいことが明らかになりました。しかし、このことのみでは、顔色変化を検出できることが3色型色覚の進化や維持にどのように関わっているかはわかりません。3色型色覚の原動力は果実採食ですが、それが進化の段階で顔色検出など、別の場面にも有効に働くようになったのかもしれません。また、果実や社会的シグナルの検出は、3色型色覚が有効である場面の例に過ぎないのかもしれません。今後も、霊長類の行動や生態学的意義と照らし合わせ、幅広く調べていく必要があります。
参考文献
Changizi MA, Zhang Q, Shimojo S. 2006 Bare skin, blood and the evolution of primate colour vision. Biology Letters, 2, 217–221. (doi:10.1098/rsbl.2006.0440)
Hiramatsu C, Melin AD, Allen WL, Dubuc C, Higham JP. 2017 Experimental evidence that primate trichromacy is well suited for detecting primate social colour signals. Proceedings of the Royal Society of London B, 20162458. (doi:10.1098/rspb.2016.2458)
この記事を書いた人

- 九州大学芸術工学研究院で視覚の研究をしています。特に動物の色覚の進化と多様性に興味を持っています。近頃は、ヒトの中での色覚の多様性に着目して研究を進めています。他者が見る世界を全く同じように体験することはできませんが、視覚の仕組みを理解することによって想像することができます。この過程において人類、また自分自身がどのような存在なのかについても客観視できることが、進化研究の面白さだと思います。



