アスパラガスのよもやま話 – 性決定遺伝子の候補を中心に
アスパラガスの生態・雌雄性
食用アスパラガス(Asparagus officinalis)は寒冷地でも春先に収穫できる貴重な野菜です。食用アスパラガスは多年生作物であり、露地栽培においては、春に萌芽し、それから初冬にかけて大きく成長して生殖と光合成を行い、根系に栄養分を移動させて地上部を枯死させ越冬し、翌年春に根から再び萌芽する、というサイクルを繰り返します。私たちが食べるのは萌芽してきた若茎ですが、商品価値のある太い若茎は、3年以上かけて栄養を蓄え太った根系からのみ、得られます。
食用アスパラガスの株には雌雄の別があります。雌株には雌花だけが着き、雄株には雄花だけが着きます。雌花においては雌しべが発達するものの雄しべは退化し、雄花においては雄しべが発達するものの雌しべは発達を停止します(下図)。アスパラガスの雌雄は、通常、このような花の形態の違いにより判別されます。露地栽培において食用アスパラガスは、発芽後1年目は花を着けないため、花の形態による雌雄の判別には長い時間を要します。
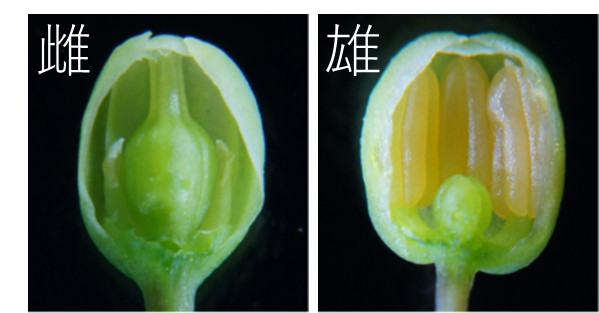
食用アスパラガスの圃場では、マルハナバチなどが受粉を媒介します。受粉が起こると、雌花の雌しべには種子が生じます。種子から生じた幼い株が、土壌中の養分をめぐって高齢な株と競合するため、種子は生産管理上疎まれます。このため、生産には雌株よりも雄株が好んで用いられます。とはいえ、雌株を圃場から排除するのは困難であり、市場に出回る若茎にも雌株由来のものが多く含まれているようです。花の性質に加えて、若茎の性状・収量や耐病性についても雌株と雄株との間で差があるという報告がありますが、これらについては未だに議論がなされています。ともあれ、食用アスパラガスの性決定の分子メカニズムを明らかにすることは、農業生産や育種の上でも重要です。
ヒトの性は性染色体XとYの組み合わせにより決定され、女性はXX、男性はXYの組み合わせを持ちます。アスパラガスの性決定様式もこれと同様であり、雌株はXX、雄株はXYの組み合わせの性染色体を持ちます。ヒトなどのX染色体とY染色体は、その長さや構造が互いに大きく異なりますが、食用アスパラガスのX染色体とY染色体は、長さや構造が殆ど同一です。ヒトなどのY染色体は全体として雄性化の機能を発揮する一方、食用アスパラガスのY染色体については、その一部のみが雄性化の機能を担っていると考えられます。すなわち、食用アスパラガスの性は比較的少数の遺伝子により決定される、ということです。雌花と雄花における雌しべと雄しべの発達の仕方を鑑みると、それらの遺伝子は、雌しべの発達を抑制し、雄しべの発達を促進するものであると推定できます。しかし、それら遺伝子の実体は長らく不明でした。
食用アスパラガスの性決定遺伝子の候補・AoMYB35の発見
モデル植物・シロイヌナズナ(Arabidopsis thaliana)においてはAtMYB35という遺伝子が失われると雄しべの発達が不全となります。先行研究において、食用アスパラガスの雌花と雄花における遺伝子産物が網羅的に解析されており、性決定遺伝子の手がかりを求めて私たちはそのデータを再解析しました。これにおいては、興味深いことに、AtMYB35に似た遺伝子の産物は、食用アスパラガスの雄花において見られるものの、雌花において全く見られませんでした。この遺伝子(AoMYB35)について解析を進めたところ、さらに興味深いことが明らかになりました。食用アスパラガスの雌株は、AoMYB35の産物だけでなく、その基となるAoMYB35の遺伝子そのものを欠いていたのです。
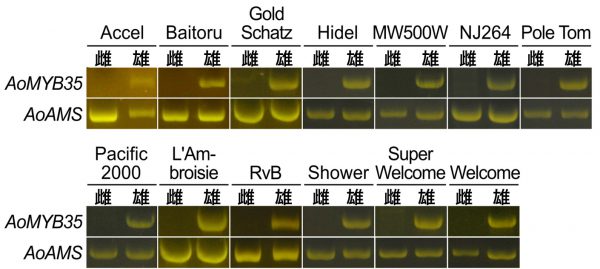
さらに解析を進めると、雄花の中でも特に発達初期の雄しべにおいて多量のAoMYB35の産物が蓄積することが明らかになりました。食用アスパラガスの雌花における雄しべの退化は、AoMYB35の欠損に起因する可能性があります。AoMYB35は食用アスパラガスの性決定遺伝子の候補として有力なものであると考えています。
今後の展望
通常、花の形態による雌雄判別は株の発芽から1年以上を要しますが、AoMYB35を遺伝子診断・DNA鑑定のような形で利用することにより、雌雄判別に要する期間を大幅に短縮することができます。このような技術は、食用アスパラガスの育種を省力化するのに役立つと考えられます。
現段階では、「AoMYB35が食用アスパラガスの性決定遺伝子である」とは言い切れません。雄花においてAoMYB35の産物は雄しべには蓄積しますが、雌しべには蓄積しません。これが雄花の雌しべの発達を抑制するということはあり得るのか? また、AtMYB35を欠損したシロイヌナズナ変異体の花においては、正常な花粉粒が作られませんが、雄しべは退化しません。これは食用アスパラガスの雌花の形態とは合致しないが、それについてはどう考えるのか? これらの問いに答えるためには、人為的に食用アスパラガス雌株にAoMYB35を導入したり、雄株のAoMYB35の機能を破壊したりする必要があると考えており、そのような実験を実際に行うべく試行錯誤しています。さらに、AoMYB35に加えて複数の遺伝子が性を決定するという可能性もあります。AoMYB35の近傍に存在する遺伝子はそのような遺伝子の候補であると考えており、それらを同定する試みも行おうとしています。これらが成功すれば、食用アスパラガスの性を転換させることが可能になるかもしれません。そのような技術も、食用アスパラガスの育種を効率化・省力化するのに貢献すると考えられます。
おまけ – 植物の雌雄性について思うこと
夏になると、食用アスパラガスの圃場にはたくさんのマルハナバチがやってきて受粉を助けます。そのような様を見ていると、AoMYB35を失うということは大した問題ではないのだろうという気にさせられます。メロン、キュウリ、マメガキなども雌雄性を持ち、それらについては性決定遺伝子が同定されていますが、それら遺伝子はすべて異なり、MYB35とも異なります。ヒロハノマンテマなど一部のマンテマ属植物も雌雄性を持ちますが、それらは形態の相異なる性染色体を有しています。交配が起こりやすい環境下では、雌しべ・雄しべの機能損失はさまざまな形で案外容易に起こり、保存され、進化するのかもしれません。雌雄性は、個体間での遺伝子の交換を促します。細胞質雄性不稔や自家不和合性(詳しくは述べません)も植物遺伝子の交換を促進する現象・性質ですが、これらと比べても雌雄性の発現パターン・メカニズムは多種多様であるように思われます。「みんなちがって みんないい」のは間違いありませんが(動物などの雌雄性もそうですが)、そこにどれほどの共通原理があるのか? 生産に活かすことはできるのか? 植物の雌雄性研究の面白さと難しさを感じつつ、その発展に僅かばかりでも貢献できればと思う今日このごろです。
参考文献
Tsugama D, Matsuyama K, Ide M, Hayashi M, Fujino K, Masuda K. (2017) A putative MYB35 ortholog is a candidate for the sex-determining genes in Asparagus officinalis. Scientific Reports 7: 41497.
この記事を書いた人
- 東京大学大学院農学生命科学研究科にて博士号を取得(実験は東京大学アジア生物資源環境研究センターにて実施)。博士課程在籍時には、日本学術振興会特別研究員(DC1)として、イネの鉄利用に関わる遺伝子について研究を行った。また、シロイヌナズナのストレス応答や細胞内信号伝達に関わる遺伝子についても研究を行った。その後、ペンシルバニア州立大学客員研究員、日本学術振興会特別研究員(PD)を経て、現職の北海道大学大学院農学研究院助教に着任。シロイヌナズナの接触刺激応答に関わるVIP1という遺伝子についてポスドク時代から継続して研究すると共に、アスパラガス等の野菜に関しても強く興味を持ち、研究を行っている。
この投稿者の最近の記事
 研究成果2017年4月27日アスパラガスのよもやま話 – 性決定遺伝子の候補を中心に
研究成果2017年4月27日アスパラガスのよもやま話 – 性決定遺伝子の候補を中心に


