受精障害はなぜおこるのか? – オスだけが持つY染色体の遺伝子の役割に迫る
Y染色体とゲノム編集
Y染色体はオスにしか存在しません。それゆえ体の大きさや生殖機能など、オスのさまざまな特徴にY染色体が関与していることが考えられます。Y染色体はX染色体に比べ非常に小さく、染色体にのっている遺伝子数も少ないのですが、実は個々の遺伝子の機能はあまりわかっていません。これは、遺伝子の機能を調べるときのゴールデンスタンダードはノックアウトマウスを作製することですが、ES細胞を使用して相同組み換えを用いる従来の方法では、Y染色体遺伝子のノックアウトマウスの作製が困難であったことが理由のひとつであると言えます。その原因は、Y染色体が反復配列を多く含むなど、特徴的な構造を取っていることなどが考えられます。それを克服したのが、近年話題になっているゲノム編集技術であるTALENやCRISPR/Cas9システムです。特にCRISPR/Cas9システムは、2012年にゲノム編集技術として報告されましたが、その簡便性から今では世界中で広く使われるようになっています。私たちの研究グループはこのゲノム編集技術をいち早く取り入れ、Y染色体上の遺伝子の機能解析を行ってきました。
Y染色体の遺伝子 Zfy1,Zfy2とは?
私たちが注目した遺伝子は、Y染色体短腕上にあるZfy1とZfy2です。この遺伝子は1980年代に性決定遺伝子として注目を集めましたが、その後Sryという遺伝子が真の性決定遺伝子であることが判明したため、しばらくのあいだ忘れ去られてしまい、その機能は長年不明でした。近年、この遺伝子が、精子形成(特に精子頭部や尾部の形態形成)に関与する報告が複数なされ、再び注目を集め始めましたが、過去の報告はいずれもマウスにZfy1やZfy2を導入する方法を取っており、ノックアウトマウスでの報告はありませんでした。そこで私たちは、CRISPR/Cas9システムを用いて、このZfy1ノックアウトマウス(Zfy1KO)、Zfy2KO、Zfy1/2ダブルノックアウトマウス(Zfy1/2DKO)を作製し、その解析を行いました。
ノックアウトマウスの解析
Zfy1とZfy2はY染色体上に存在し、生殖機能に関与している可能性が高いため、まずノックアウトマウスをメスと交配させその妊孕性を調べました。するとZfy1KOと交配させたメスのマウスは、問題なく妊娠することができるのに対し、Zfy2KOと交配させたメスのマウスは妊娠しにくく、Zfy1/2DKOでは不妊となりました。この原因を調べていくと、Zfy2KOやZfy1/2DKOが持つ精子は、下図のように頭部や尾部に奇形があり、運動性も落ちていることがわかりました。またその程度は、Zfy2KOよりもZfy1/2DKOのほうが重度であり、Zfy1とZfy2では、Zfy2のほうが役割としては重要ですが、互いに機能を補い合っていることが考えられました。
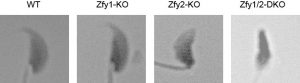
Zfy1/2DKOの精子は、卵子に精子を直接ふりかける体外受精でも受精できませんでした。そもそも精巣上体に貯留している精子は、そのままでは受精することができません。メスの生殖器官を通過する過程で受精能を獲得し、卵周囲の顆粒膜細胞に反応して先体反応が起こり、最終的に精子と卵子の細胞膜が癒合し受精が起こります。Zfy1/2DKOの精子では、受精能獲得や先体反応が起こらないことがわかりました。
こんなだめだめなZfy1/2DKO精子ですが、精子を卵子に直接注入する顕微授精を行って、はじめて受精することができました。しかし、その受精効率は非常に低く、また受精しても多くの胚が早い段階で発生を停止してしまいました。この顕微授精における受精効率の低さを調べていくと、卵子の活性化が起こっていないことがわかりました。通常、第二減数分裂中期で減数分裂が止まっている卵子は、受精時に精子由来の卵活性化因子により、減数分裂が再開します。つまり、Zfy1/2DKO精子は卵の活性化に障害があり、顕微授精でも受精効率が低下していると考えました。また、わずかに受精しても発生が早い時期に止まる原因を調べていくと、精子由来の染色体の断片化が起きていることがわかりました。
Zfy, Zfy2の役割を明らかに
では、Zfy1, Zfy2はどのような役割を持っているのでしょうか? Zfy1, Zfy2は、Zinc fingerというDNAに結合することができるモチーフを複数持っており、その構造から転写因子と考えられています。転写因子とは、ほかの遺伝子の発現の制御に関わる遺伝子のことです。しかし、Zfy1とZfy2が制御している遺伝子に関しては、全くわかっていません。それゆえZfy1/2DKOの精子では、さまざまな異常が観察されましたが、これらの異常はZfy1, Zfy2が直接関与しているのではなく、ほかの遺伝子の発現が上昇または低下しているために起こっているものと考えました。そこで野生型(正常)マウスとZfy1/2DKOの精子を用いて、タンパク質を網羅的に調べる質量分析を行いました。その結果、Zfy1/2DKOの精子は多くのタンパク質の発現が、野生型の精子と比べて低下していることがわかりました。そしてそのなかにZfy1/2DKOの表現型を説明しうるタンパク質が複数含まれていました。それは卵活性因子であるPlcz1、先体反応に関わるPrss21, Plcd4、精子頭部の形態形成に関わるHttです。つまりZfy1/2DKOで観察されたさまざまな異常の少なくとも一部は、これらの遺伝子の発現が低下していることで起こっている可能性があります。
最後に
今日不妊治療の現場において体外受精や顕微授精は広く行われていますが、顕微授精を用いたとしても受精できない、または受精したとしてもその後の発生があまり良くない、という患者さんは少なからず存在し、治療に難渋します。このZfy1,Zfy2のさらなる機能解明が、そのような方への治療の一助となることを期待し、今後も研究を続けていきたいと思います。
参考文献
Nakasuji, T., et al., Complementary Critical Functions of Zfy1 and Zfy2 in Mouse Spermatogenesis and Reproduction. PLoS Genet, 2017. 13(1): p. e1006578.
この記事を書いた人

-
中筋貴史(なかすじたかし)(写真左)
東京医科歯科大学医歯学総合研究科生殖機能協関学、大学院生。東京医科歯科大学医学部医学科を卒業後、初期臨床研修を経て産婦人科医として勤務。2013年4月より現職。産婦人科の中でも不妊治療に興味を持っていることから大学院の研究テーマとしてY染色体と精子形成に関して浅原研究室で共同研究を行っている。今後はZFYと男性不妊症との関連を明らかにしていきたいと考えている。
淺原弘嗣(あさはらひろし)(写真右)
東京医科歯科大学医学部、システム発生・再生医学分野、教授。岡山大学医学部卒業、整形外科医のトレーニングの後、ハーバード大学医学部、ソーク研究所を経て、2002年、スクリプス研究所にラボを開設。その後、国立成育医療研究センターを経て、2011年より現職。RNA階層を包括した遺伝子発現で調律される生命のレゾンデートルを、御茶ノ水の天空のラボで
一緒に追いかけませんか?ラボのホームページはこちら



