植物が緑になるか否かはどう決まる? – 根で葉緑体の発達をコントロールするしくみ
動物と植物の違いは?
動物と植物の違いはなんでしょうか? 小さな子どもに説明するなら、動き回ってなにか食べるのが動物、じっとしていて食べないのが植物、ということになるでしょう。ただ、サンゴのように動かない動物もいれば、ハエトリグサのように虫を食べる植物もいるので、例外もたくさん存在します。そこで、より詳しく説明するなら、基本的な細胞のつくりが違う、と答えることができます。動物と植物の細胞には細胞壁の有無などさまざまな相違点がありますが、なかでも特徴的なのが、色素体とよばれる細胞内小器官を持つ(植物)か、持たない(動物)か、という違いです。色素体は藻類を含む植物に特有の細胞内小器官の総称で、光合成を行う葉緑体も色素体の一形態です。色素体を持つか否かが植物と動物の生き方を決定的に違えており、色素体(葉緑体)を持つ植物は光合成によりエネルギーを確保できるので、動き回って食べ物を探す必要がないのです。
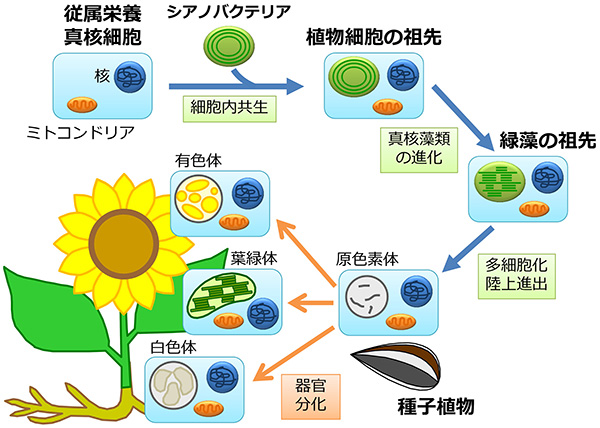
では、この違い“色素体を持つか否か”はどのように生じたのでしょうか? さまざまな証拠から、遥か昔に植物細胞の祖先が光合成を行う細菌(シアノバクテリア)を取り込み、色素体として使うようになった、と考えられています(細胞内共生説)。共生した細菌に光合成を行う能力があった、というのが最も重要な点で、それにより植物細胞は光と水という非常にありふれた材料からエネルギーを取り出すことが可能になり、地球の環境を変えるほどの発展を遂げたのです。
色素体は細胞の機能に応じてさまざまに形を変える
このような進化的な背景を考えると、色素体の原型は光合成能力を持つ葉緑体であるといえます。実際、単細胞藻類などでは、色素体はもっぱら葉緑体として存在し、盛んに光合成を行うことで細胞の増殖を助けます。一方、多細胞の高等植物では、色素体は葉緑体以外にも、細胞の役割に応じてさまざまな機能を持った形態をとります。たとえば、大根の根の細胞では無色の白色体が、トマトの果実の細胞ではカロテノイドを持つ有色体が、ジャガイモの塊茎の細胞ではデンプン粒を貯め込んだアミロプラストが発達します。葉緑体も含め、これらの多様な色素体は原色素体とよばれる未分化な色素体から分化するほか、発達過程や生育環境に応じて相互に変換することも知られています。トマトの例でいうと、果実が熟すにつれて緑色から赤色に変わるのは、緑色の葉緑体が赤いリコペンを含んだ有色体に変身するからです。植物は多様な環境に適応するために、進化の過程で葉緑体をさまざまな機能を持つ色素体へと分化させるようになったのでしょう。
色素体の分化を制御するしくみは?
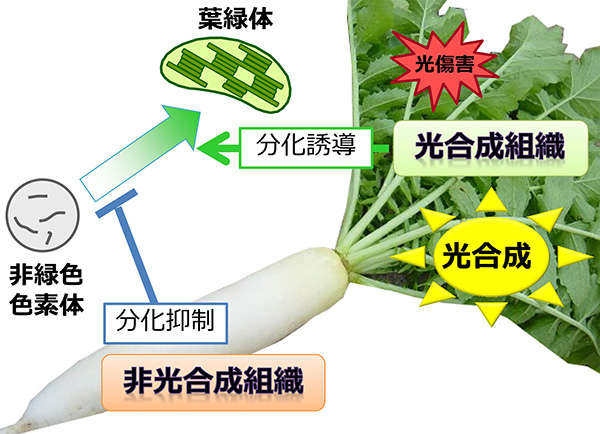
このように、色素体の分化が植物の生き方を決めていると言っても過言ではありませんが、色素体の分化を調節するしくみについては、実のところまだよくわかっていません。特に、葉緑体が担う光合成は諸刃の剣で、吸収した光エネルギーを反応にうまく使えないと、生じた活性酸素により細胞がダメージを受けます。そのため、植物は組織や環境に応じて葉緑体の発達を厳密にコントロールする必要がありますが、葉緑体の分化が細胞ごとにどのように決められるのかについては、ほとんど明らかになっていません。これまでの葉緑体分化の研究は、光合成組織でどのように葉緑体が発達するのか、という「正の制御」に関するものがほとんどで、非光合成組織ではなぜ葉緑体が発達しないのか、という「負の制御」に関してはあまり注目されていませんでした。しかし、葉緑体が色素体の原型であると考えると、非光合成組織ではむやみに葉緑体が分化しないように強く抑制しているともいえます。そこで私は、非光合成組織で葉緑体の発達を抑制するしくみを明らかにできれば、色素体の分化制御機構の解明に近づけると考え、モデル植物であるシロイヌナズナの根を材料に研究を開始しました。
植物ホルモンのバランスが葉緑体の分化を左右する
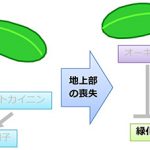
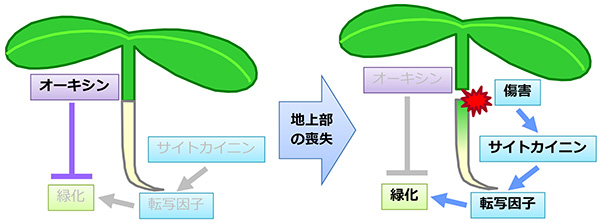
一般に、植物の根は成長に必要な炭素源を光合成器官である葉に依存しています。自らは光合成をする必要はないので、根は葉緑体をほとんど発達させません。実際、シロイヌナズナの根に光を当てても、葉緑体の緑色の色素であるクロロフィル(葉緑素)はほとんど作られません。しかし、光合成組織である地上部(葉や茎)を切り取ると、根でクロロフィルの合成が活性化し、葉緑体も増えることに気づきました。このことは、通常は光合成を行わない根の細胞も、環境に応じて葉緑体を発達させる能力を持つことを意味します。そこで、この現象を詳しく解析した結果、植物ホルモンのオーキシンが根における葉緑体の発達を抑制することを突き止めました。地上部から切り離した根をオーキシンを含む培地で生育させたところ、クロロフィルの蓄積が抑えられました。反対に、地上部から根へのオーキシン輸送やオーキシンの情報伝達が弱くなった変異体では、地上部を切らずとも根が緑色になりました。これらの結果から、通常は、根の葉緑体発達は地上部から輸送されるオーキシンの作用によって抑制されていますが、地上部を失ったときにはオーキシンによる抑制がなくなり、根で葉緑体が発達しやすくなることが明らかとなりました。
さらに、サイトカイニンという別の植物ホルモンが、オーキシンとは反対に根での葉緑体の発達を促進することを見出しました。サイトカイニンを与えると根のクロロフィル量が増加するのに加え、地上部を切り離した根ではサイトカイニンに対する応答が強くなることもわかりました。このサイトカイニン応答は根の緑化に必須であり、サイトカイニン応答因子(BタイプARR)の変異体では、地上部を切除した際の根の緑化応答がまったく見られなくなりました。また、一連の反応には傷害応答因子WIND1が深く関与しており、WIND1の機能を抑制した植物体ではサイトカイニン応答が弱くなるとともに、葉緑体の発達も抑制されることがわかりました。以上の結果から、通常、根の細胞ではオーキシンの作用により葉緑体の発達が抑制されていますが、地上部を失った際にはオーキシンによる抑制が解除され、それと同時にWIND1を介してサイトカイニン応答が活性化され、根での葉緑体の発達が促進されることが明らかになりました。
転写因子の活性化で根が緑に
最後に、植物ホルモンの下流で働く因子にも簡単に言及したいと思います。私は一連の研究から、オーキシンとサイトカイニンによる葉緑体の分化制御には複数の転写因子(DNAに結合し遺伝子の転写を調節するタンパク質)が関わっていることを突き止めました。実際、そのような因子を人工的に過剰に作らせると、葉緑体の発達が誘導されることで白い根が緑になり、光合成能力も向上することが確認できました。つまり、転写因子を人為的にコントロールすることで、本来光合成をしない根でも葉緑体を作らせることができたのです。この技術をさらに発展させれば、将来的には植物の葉緑体の分化を人工的にコントロールできるようになるかもしれません。
おわりに
エネルギーの供給源である地上部を失うことは、植物にとって最大のピンチです。そのような危機に対し、植物は残った根で葉緑体を発達させ光合成を行うことで、生き残る可能性を少しでも高めていると考えられます。草むしりで根を残してしまうと、そこからまた葉が出てくることがありますが、以上のような葉緑体分化のしくみが働いているのかもしれません。今後さらに研究を進めて、色素体の分化を制御するしくみと、それに基づく植物の多様な生き方を明らかにしたいと考えています。
参考文献
Kobayashi, K., Ohnishi, A., Sasaki, D., Fujii, S., Iwase, A., Sugimoto, K., Masuda, T. and Wada, H. (2017) Shoot removal induces chloroplast development in roots via cytokinin signaling. Plant Physiol. doi: 10.1104/pp.16.01368.
Kobayashi, K., Sasaki, D., Noguchi, K., Fujinuma, D., Komatsum, H., Kobayashi, M., Sato, M., Toyooka, K., Sugimoto, K., Niyogi, K. K., Wada, H. and Masuda, T. (2013) Photosynthesis of root chloroplasts developed in Arabidopsis lines overexpressing GOLDEN2-LIKE transcription factors. Plant Cell Physiol. 54: 1365-1377.
Kobayashi, K., Baba, S., Obayashi, T., Keränen, M., Aro, E.M., Fukaki, H., Ohta, H., Sugimoto, K. and Masuda, T. (2012) Regulation of root greening by light and auxin/cytokinin signaling in Arabidopsis. Plant Cell 24: 1081-1095.
この記事を書いた人

この投稿者の最近の記事