ミトコンドリアの健康管理には、他の細胞小器官との連携が重要
ミトコンドリアを狙って壊すシステム「マイトファジー」
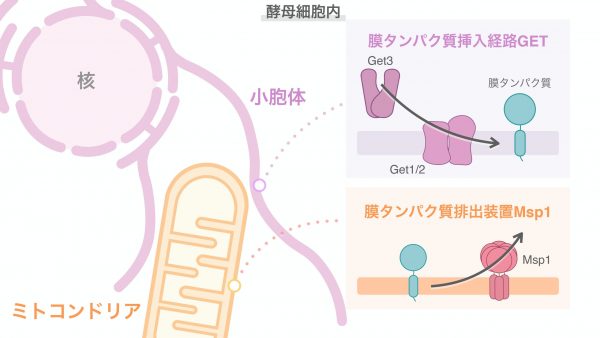
真核生物の細胞の中には膜で囲まれたいくつかの区画があり、それぞれの区画は細胞小器官(オルガネラ)と呼ばれ、細胞機能を分担しています。そのなかでも、私たちが研究しているミトコンドリアは外膜と内膜の二重の膜からなる細胞小器官で、主に細胞活動に必要なATPを供給する細胞の「発電所」です。そのため細胞は、エネルギー需要の変動に応じてミトコンドリアの量を増減させる必要があります。またミトコンドリアは、酸化ストレスの過剰な蓄積によりダメージを受け、細胞にとって有害となりうるため、品質の低下したミトコンドリアは迅速かつ適切に分解される必要があります。ミトコンドリア特異的オートファジー「マイトファジー」は、損傷したり余分に生じたりしたミトコンドリアを分解することで、ミトコンドリアの品質や量を調節する機構のひとつです。細胞を普段私たちが過ごしている部屋にたとえるなら、部屋の中に溜まってきたゴミや使わなくなった家具を捨てたりして整理する仕組みと同じようなことが、細胞の中でも起こっているのです。そうした整理整頓を怠ると生活に支障をきたすのと同じように、マイトファジーが正常に起きなくなると細胞機能に多様な異常を引き起こすことが報告されており、神経変性疾患やがんといった疾患につながる可能性も示唆されています。今回私たちは、マイトファジーを介したミトコンドリアの分解機構に、小胞体の膜タンパク質挿入経路GETと、ミトコンドリアの膜タンパク質排出装置Msp1が関わることを明らかにしました。
私たちは、「そもそもマイトファジーはどのように起きているのか?」という点を明らかにすべく、モデル生物である出芽酵母を使って研究を進めてきました。出芽酵母においてマイトファジーが誘導されるとき、他の細胞質成分と区別してミトコンドリアのみを分解の標的とすべく、その目印として働くタンパク質Atg32がミトコンドリア外膜に蓄積します。ミトコンドリア上に集積したAtg32は、オートファジーの進行に重要な役割を果たすAtg8やAtg11などのタンパク質と結合し、マイトファジーを開始する複合体を形成します。
私たちの研究室では、出芽酵母マイトファジーの分子機構の解明を目指してゲノムワイドな遺伝子スクリーニングを行い、マイトファジーに重要だと考えられる遺伝子を複数同定してきました。その過程で見出されたのが、膜タンパク質を小胞体膜へ挿入する機能を持つGET経路の構成因子(Get因子)です。小胞体も細胞小器官の一種で、細胞内を張り巡るネットワーク構造をとっており、脂質やステロイドの合成、タンパク質の折りたたみや成熟化などを担っています。GET経路の中心となる因子は小胞体膜に存在することがわかっていましたが、小胞体上の因子が別種の細胞小器官であるミトコンドリアの分解にどのように関わるかはまったくの未知でした。
タンパク質配送装置の異常が招くマイトファジーの不具合
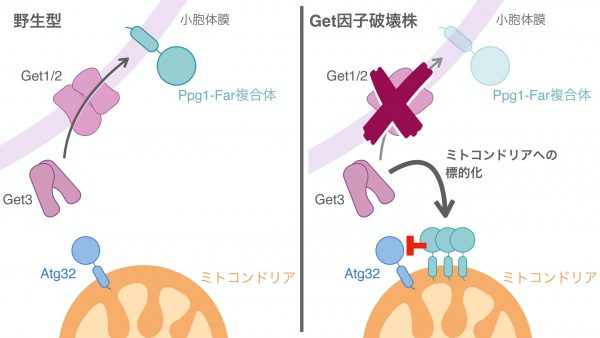
この疑問を解決すべく、まずはGet因子を欠損した酵母株を使ってマイトファジーのどのステップに異常が起きているかを観察しました。するとGet因子の欠損株では、マイトファジー誘導条件におけるAtg32のリン酸化レベルが減少していることを突き止めました。リン酸化を介したAtg32の活性化は、Atg32とAtg11の結合を安定化させ、マイトファジーを駆動させるのに重要です。この知見と一致するように、Get欠損株ではAtg32-Atg11相互作用が不安定化していることもわかりました。それでは、Get因子欠損時のAtg32のリン酸化レベルの減少は、どのようにして起こるのでしょうか。
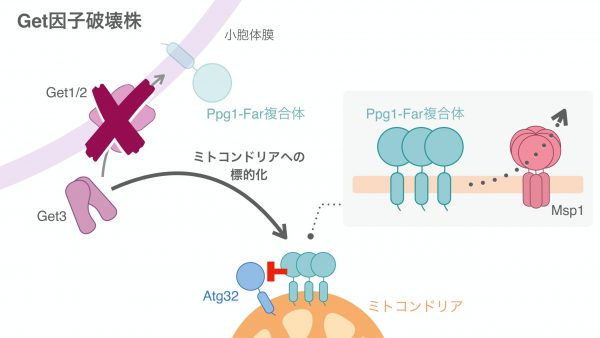
ここで、Atg32の脱リン酸化を担い、Atg32-Atg11相互作用の抑制に寄与するタンパク質として知られていたPpg1に着目しました。Ppg1は小胞体膜タンパク質Farと結合し、主に小胞体に存在することが示唆されていましたが、Get欠損株ではFarタンパク質は小胞体局在を失い、ミトコンドリアの周りを綺麗に囲うような局在を示す様子が観察されました。この観察結果から、GET経路はPpg1-Far複合体を小胞体に繋ぎ止めるのに重要であること、そしてGet欠損株ではPpg1-Far複合体が過剰にミトコンドリアへと局在することで、Atg32のリン酸化を抑えてしまうと考えました。この可能性を検証するため、Get欠損株でFarタンパク質だけを人工的に小胞体に局在させることでミトコンドリアへの局在を解消したところ、低下していたAtg32-Atg11相互作用、マイトファジーが有意に回復することを見出しました。この実験を行うとき、期待した結果になるかどうかわからずいつになく緊張していましたが、これらの結果は、GET経路がPpg1-Far複合体を小胞体膜へ繋留しミトコンドリアへの局在を防ぐことで、Atg32のリン酸化とマイトファジーの駆動に寄与することを示唆しています。
Get欠損株では、種々の小胞体膜タンパク質がミトコンドリア膜へと配送されることが知られています。ミトコンドリアには、こうした普段はほとんど配送されない「よそ者」タンパク質をきちんとミトコンドリア膜から取り除く装置が存在しています。それが、ミトコンドリア外膜タンパク質Msp1です。興味深いことに、Get因子とMsp1を二重欠損させてみたところ、マイトファジーの効率が相乗的に減少することがわかりました。この際、Farタンパク質を人工的に小胞体に局在させると、マイトファジーが野生型と同程度まで回復しました。これらの結果から、Msp1がミトコンドリア外膜上のPpg1-Far複合体を取り除くことで、マイトファジーの進行に働いている可能性が考えられます。
お隣さん同士のオルガネラの連携が、ミトコンドリアの品質管理に重要
「マイトファジーを駆動する因子はどのように活性化されるのか?」という点について、小胞体などの異種オルガネラに存在する因子がどのように関わるかはほとんど不明でした。そうしたなか、本研究では、小胞体の膜タンパク質挿入経路がマイトファジー駆動因子の活性化機構に寄与しているという証拠を得、さらにミトコンドリアの膜タンパク質排出装置と絡めて、マイトファジー制御の一端を明らかにすることができました。こうした視点からマイトファジーの仕組みを解明することは、オルガネラ間連携の新たな役割の理解にもつながり、細胞生物学の分野に広く影響を与えるものと期待しています。
参考文献
- The GET pathway serves to activate Atg32-mediated mitophagy by ER targeting of the Ppg1-Far complex
Mashun Onishi, Mitsutaka Kubota, Lan Duan, Yuan Tian, Koji Okamoto
Life Science Alliance 6(4):e2022016402023 2023年1月25日 - Molecular mechanisms and physiological functions of mitophagy
Mashun Onishi, Koji Yamano, Miyuki Sato, Noriyuki Matsuda, Koji Okamoto
The EMBO Journal 40(3):e104705 2021年2月1日 - The ER membrane insertase Get1/2 is required for efficient mitophagy in yeast
Mashun Onishi, Sachiyo Nagumo, Shohei Iwashita, Koji Okamoto
Biochemical and Biophysical Research Communications 503(1):14-20 2018年9月3日
この記事を書いた人

-
大西真駿(写真左)
マックス・プランク老化生物学研究所(MPI-AGE, ドイツ)・ポストドクトラルフェロー。大阪大学大学院生命機能研究科のミトコンドリア動態学研究室(岡本浩二准教授)のもとで博士号(理学)を取得。その後、同研究室にて日本学術振興会特別研究員、大阪大学特任研究員を経て、2021年10月よりドイツに渡航・現職。現在はMPI-AGE のThomas Langer教授が主宰する研究室で、「ミトコンドリアが細胞の生と死を決める分子メカニズム」を研究している。
岡本浩二(写真右)
大阪大学大学院生命機能研究科ミトコンドリア動態学研究室(准教授・PI)。広島大学大学院理学研究科修了・博士(理学)(1995年3月)。東北大学遺伝生態研究センター・大瀧保研究室、テキサス大学サウスウェスタンメディカルセンター・Ronald Butow研究室、ミュンヘン大学生理化学研究所・Walter Neupert研究室、ユタ大学医学部生化学科・Janet Shaw研究室、基礎生物学研究所/東京工業大学先進研究院・大隅良典研究室を経て、2010年4月より現職。現在の研究テーマは、「ミトコンドリアの質・量管理システムの分子機構と生理機能の解明」。
この投稿者の最近の記事
 研究成果2023年2月16日ミトコンドリアの健康管理には、他の細胞小器官との連携が重要
研究成果2023年2月16日ミトコンドリアの健康管理には、他の細胞小器官との連携が重要


