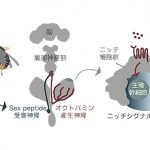
神経が幹細胞の分裂を活性化する仕組み – 交尾によって生殖幹細胞が増加するメカニズムに迫る
マクロからミクロ − 環境情報から幹細胞制御までのつながり
皆さんは、ニュースなどで「幹細胞」という言葉を聞いたことがあるでしょうか。幹細胞とは自己増殖能力や、異なる性質を持つ細胞へと分化する能力を持っている細胞で、わたしたちの体にもたくさん存在しています。体毛が抜けても新たな毛が生え、転んでケガをしても皮膚の傷が治るのは幹細胞が増殖・分化して再生するためです。
この幹細胞の増殖・分化の刮目すべきところは、「必要な時に必要な分だけ増殖・分化すること」にあります。たとえば、ケガをしていないのに皮膚の幹細胞が過剰に増殖・分化を始めてしまうと、皮膚には過剰な細胞の塊が形成されてしまいます。では、幹細胞の増殖・分化は一体どのようにして適切に制御されているのでしょうか?
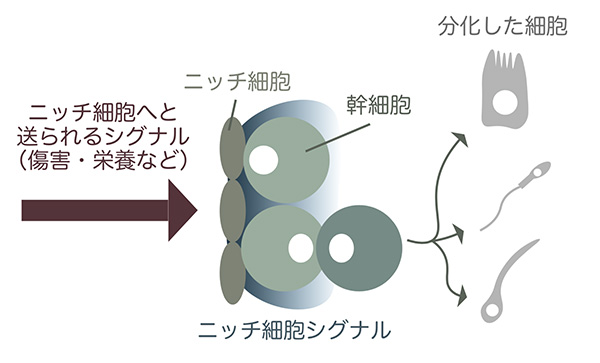
これまでの多くの研究から、幹細胞の増殖・分化は、幹細胞の近くにある細胞たちによって調節されていることが明らかにされています。その細胞たちは”ニッチ細胞”と呼ばれ、幹細胞に増殖や分化を調節するためのシグナルを送っています。いわばニッチ細胞は幹細胞の増殖や分化を調節する司令塔のような存在で、生物が生存するための組織の恒常性の維持に一役かっているのです。
ニッチ細胞の概念はRay Schofieldの造血幹細胞の研究の過程で提唱されましたが、それ以降、さまざまな動物の多種多様な幹細胞においてニッチ細胞の存在と幹細胞の増殖と分化における重要性が支持されてきました。
しかし、ニッチ細胞の意義が受け入れられるにつれ、今度は新たな疑問が浮かび上がります。一体ニッチ細胞はどのようにして脱毛やケガといった、幹細胞による回復を必要とする状態を感知しているのでしょうか。その感知には、ニッチ細胞へと情報を伝達する、さらに上流の者たちの存在が必要不可欠です。幹細胞とニッチ細胞のシステムによりますが、ときにはニッチ細胞へと伸びる神経が、またあるときはニッチ細胞と密接している別のタイプの細胞が、ニッチ細胞へと”幹細胞の分裂を活性化せよ!”という情報を送るのです。
近年、このニッチ細胞の上流の因子を介した、環境からのシグナルに応じた幹細胞の分裂制御に関心が高まっています。わたしたち、ヒトをはじめとする生物は、環境というマクロな情報を幹細胞というミクロなものに伝達することで、その環境に適応して生きるための組織の恒常性を維持しているのです。
ショウジョウバエの研究から見えてきた幹細胞の動態制御の仕組み
わたしたちが主に使用している実験動物はキイロショウジョウバエ(Drosophila melanogaster)です。驚かれる方も多いかもしれませんが、キイロショウジョウバエはこの幹細胞とニッチの関係を深く追究するうえで、これまで重要な貢献をしてきました。
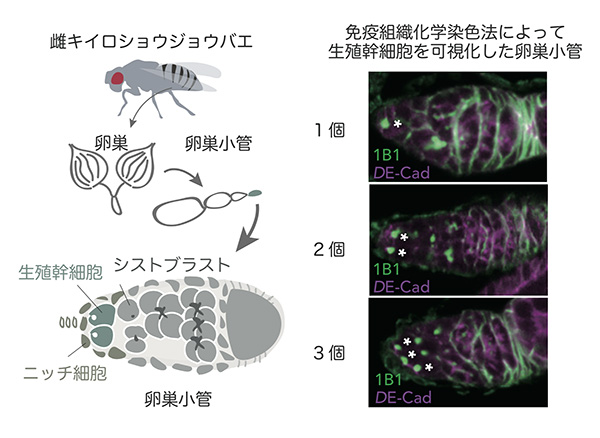
歴史的金字塔のひとつは、1998年にTing XieとAllan C. Spradlingがキイロショウジョウバエの卵をつくるもととなる生殖幹細胞のニッチ細胞を発見したことです。この発見こそが、幹細胞に接する体細胞が幹細胞の分裂を制御すること、すなわちニッチ細胞が確かに存在することを初めて特定したものでした。
その後、キイロショウジョウバエの生殖幹細胞とニッチ細胞の系を用いた多くの研究により、幹細胞とニッチの分子的、構造的な基盤の理解が加速していきました。そして、上述のような、環境からの情報による生殖幹細胞の増殖・分化の調節のメカニズムに迫る報告も数多く現れ始めました。
わたしの現在所属している研究室では、2016年に”雌キイロショウジョウバエの生殖幹細胞が、雄と交尾することによって増える”ことを報告しました。キイロショウジョウバエは、交尾後わずか24時間で30-60個の卵を産むようになり、雌1個体の産卵数は生涯を通じて300個ほどにもなります。雌はこの産卵を下支えするために、生殖幹細胞からの卵の発育を促進する必要があります。交尾後の生殖幹細胞の増加を抑えると、産卵数も減少することから、わたしたちは”交尾後の生殖幹細胞の増加は卵の発育を活性化し、産卵数を増やすために重要な現象である”と考えています。しかし、交尾の刺激がどのようにして生殖幹細胞の分裂を制御するニッチ細胞へと送られるのか、という疑問が残されていました。
神経伝達物質オクトパミンが交尾後の生殖幹細胞増加を制御
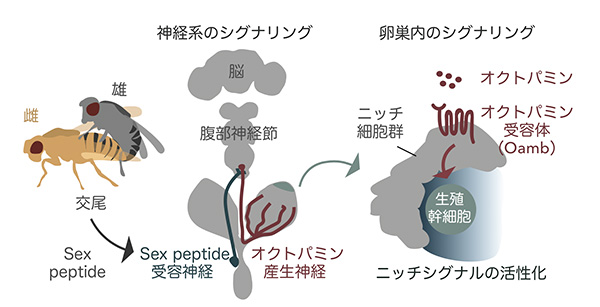
そこでわたしたちは、交尾の刺激をニッチ細胞で受け取って、生殖幹細胞の増加を誘導する因子を探索しました。その結果、昆虫の神経伝達物質であるオクトパミンと、オクトパミンを受け取るオクトパミン受容体(Oamb: Octopamine receptor in mushroom body)が交尾後の生殖幹細胞の増加に必須であることがわかりました。
オクトパミンは哺乳類のノルアドレナリンの類縁物質で、動物種を越えて広く保存されています。オクトパミン受容体Oambの機能を卵巣で阻害した雌ショウジョウバエでは、交尾後に見られるはずの生殖幹細胞の増加が起こらなくなっていました。また、オクトパミンの生合成に必要な遺伝子の突然変異株においても、生殖幹細胞の増殖は交尾後に生じなくなりました。
次の疑問は、オクトパミンがどこかからやってくるのかでした。その候補としてわたしたちは、卵巣へと直接伸びているオクトパミン産生神経に着目しました。実は雌ショウジョウバエの卵巣には、オクトパミン産生神経が伸びており、その神経が産卵などの行動を調節していることが報告されていました。
そこで、わたしたちがオクトパミン産生神経を強制的に活性化したところ、雌ショウジョウバエは交尾をせずとも生殖幹細胞を増加させ、逆にオクトパミン産生神経を不活化させた際には、交尾をしても雌ショウジョウバエの生殖幹細胞の数は増えませんでした。これは、生殖幹細胞という卵をつくるもととなる幹細胞の増加が特定の神経のはたらきによって直接制御されているという、大きな証拠になりました。
生殖幹細胞の増加に重要な、交尾の情報を伝達する神経系のリレー
しかし依然として、交尾の情報がどのようにして卵巣へと伸びるオクトパミン産生神経へと伝えられるのかという疑問が残っています。実は2016年のわたしたちの報告で、オスショウジョウバエの精液中に含まれるSex peptideというタンパク質が、交尾後の生殖幹細胞の増加に重要であることを見出していました。
このオスの精液中のSex peptideは、雌の子宮付近にある神経によって受け取られ、その情報は脳へと送られることが知られています。そこで、わたしたちがこのSex peptide受容神経と卵巣へと伸びるオクトパミン産生神経の関わりを調べたところ、驚くべきことに腹部神経節(脳の一部)内でSex peptide受容神経とオクトパミン産生神経が直接的にシナプスを形成していることがわかりました。
これにより、交尾―オス精液中のSex peptide―Sex peptide受容神経―オクトパミン産生神経―生殖幹細胞ニッチという、交尾から生殖幹細胞制御までを繋ぐ1本のリレーが明らかになりました。
昆虫と哺乳類、意外な共通性
ここまでで、キイロショウジョウバエにおける交尾と生殖幹細胞の関係について触れてきました。生殖幹細胞とそれを支持するニッチ細胞は、ショウジョウバエのみならず、線虫などの多くの無脊椎動物に存在することがわかっています。
しかし実は、わたしたちヒトをはじめとする哺乳動物の多くにおいて、メス個体における生殖幹細胞の存在は否定されています。代わりに、多くの哺乳動物では未成熟な卵が卵巣内で蓄えられており、成熟とともに排卵が起こります。実は哺乳動物には、ヒトのように周期的に排卵が起こる種と、交尾によって排卵が誘導される種がありますが、そのどちらの排卵過程においても、ホルモンや神経系からの刺激が重要であると考えられています。
キイロショウジョウバエのオクトパミンに近い分子が、哺乳類のノルアドレナリンであると上述しましたが、興味深いことに、哺乳類においてもノルアドレナリン神経が排卵の誘導に重要であるという報告があります。キイロショウジョウバエと哺乳動物は見た目や体の構造は大きく異なりますが、分子レベルで見てみると、環境からの情報を生殖へとインプットするメカニズムには共通点があるのかもしれません。
さらに近年の研究で、哺乳動物においてノルアドレナリン神経が毛髪をつくる毛包幹細胞の活動を調節していることがわかっています。動物種を越えてノルアドレナリンやオクトパミンといった神経伝達物質が、”幹細胞の分裂の制御”という個体の組織の恒常性を保つうえで重要な役割を担っているようです。
キイロショウジョウバエやマウスなどのモデル生物を使用した研究により、幹細胞の動態制御の動物間での違いや共通性が浮き彫りとなり、”環境というマクロな情報を幹細胞というミクロなものにどのように伝達し、組織の恒常性を保っているのか”という大きな疑問に答える一助になるとわたしたちは考えています。
参考文献
- Spradling A, Fuller MT, Braun RE, Yoshida S. 2011. “Germline stem cells.” Cold Spring Harb Perspect Biol 3:a002642. doi:10.1101/cshperspect.a002642
- Ameku T, Niwa R. 2016. “Mating-Induced Increase in Germline Stem Cells via the Neuroendocrine System in Female Drosophila.” PLOS Genet 12:e1006123. doi:10.1371/journal.pgen.1006123
- Yoshinari Y, Ameku T, Kondo S, Tanimoto H, Kuraishi T, Shimada-Niwa Y, Niwa R. “Neuronal octopamine signaling regulates mating-induced germline stem cell increase in female Drosophila melanogaster.” eLife 9:e57101. doi: 10.7554/eLife.57101.
この記事を書いた人

-
吉成 祐人(写真左)
筑波大学生命環境科学研究科生物科学専攻在学中。交尾、栄養といった外部環境からのシグナルに生物がどのようにして適応しているのかを、そのメカニズムをモデル生物であるキイロショウジョウバエを用いて探索しています。趣味は陸上競技(長距離)。
丹羽 隆介(写真右)
筑波大学 生存ダイナミクス研究センター(TARA)教授。
2002年 京都大学大学院理学研究科修了。日本学術振興会特別研究員SPD(東京大学)およびヒューマンフロンティアサイエンスプログラム長期フェロー(Yale大学)でのポスドクを経て、2008年に助教として筑波大学に着任。2012年 同准教授。2019年 同教授。ショウジョウバエと寄生蜂を主材料として、神経とホルモンを介した器官と器官の相互作用が生命のホメオスタシスとトランジスタシスの調節に果たす役割を研究しています。