シロイヌナズナの花粉数を制御する遺伝子 – 少ない花粉と生育メリットを両立するカギ
植物の「花粉数」はどうやって決まる?
みなさんが「花粉数」と聞くと何を思い浮かべるでしょうか? 花粉症の方は真っ先に気象予報で報道される花粉飛散予想を思い出すかもしれません。花粉症の患者さんにとってはアレルギー対象となる花粉の飛散量はとても重要ですが、植物にとっても花粉数は生存戦略や進化を考えるうえで非常に重要な形質です。
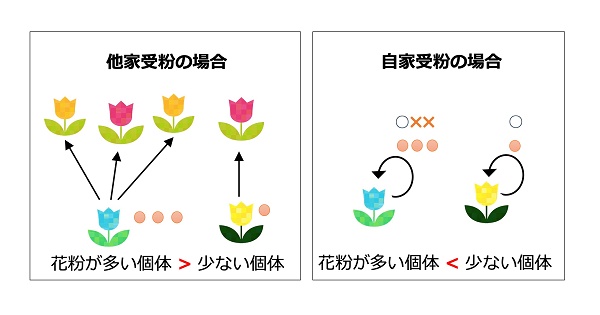
花を咲かせる植物(顕花植物)の多くは他個体と交配を行う他家受粉によって子孫を残しています。これら他家受粉の植物にとってはより多くの花粉を生産することがより多くの自分の子孫を残すことに繋がるため、多くの花粉をつくる個体が生存戦略上より有利な個体ということになります。
一方で他個体の花粉を必要とせず、自分の花粉で子孫を残すことができる自家受粉が可能な植物もいます。自家受粉を行う植物にとっては大量の花粉の生産は投資コストの無駄になると考えられ、実際に近縁種の他家受粉の植物と比較すると自家受粉を行う植物の花粉数は少ない傾向があることが知られています。
では花粉数はどのような遺伝子によって決定されているのでしょうか? 花粉数が植物によって異なることは19世紀にチャールズ・ダーウィンが論じて以来、研究者が興味を持って研究を行ってきましたが、花粉数を制御する遺伝子は明らかにされてきませんでした。
花粉数を制御する遺伝子RDP1を発見
これまでは、顕微鏡下で花粉を観察し、数えた結果を用いて1花あたりの花粉を推定していました。この方法では顕微鏡観察や計数に時間がかかるうえに、一度に計数できる花粉数が100程度と少ないことから測定誤差が大きいことも問題でした。
そこで私たちは、顕微鏡のかわりにセルカウンターを使う方法を新たに開発しました。セルカウンターは1回の解析で約2万粒までの花粉を1分以内に計測することができるため、この方法を用いることで迅速に多数の花粉を計測することができるようになりました。
自家受粉を行うモデル植物であるシロイヌナズナ144系統の花粉数を計測したところ、花粉数の少ない系統では1花あたり2,000粒、花粉数が多い系統では8,000粒程度の花粉を生産することがわかりました。この情報をもとにゲノムワイド関連解析(Genome-wide association study)を行い、花粉数に関わる遺伝子領域の特定を試みました。ゲノムワイド関連解析は、系統ごとのゲノム配列と形質(今回は花粉数)を比較することで、興味のある形質に関わる遺伝子にピンポイントで迫ることができる手法です。
解析の結果、第1染色体上に最も花粉数と関連度が高い領域が存在することがわかり、その領域内には3つの遺伝子が存在することがわかりました。この3遺伝子それぞれについて、ゲノム編集手法のひとつであるCRISPR/Cas9を用いて遺伝子の機能を破壊する実験を行いました。
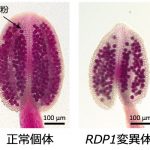
このうちひとつは翻訳・リボソーム制御に関わる遺伝子と相同性を示す新規遺伝子で、その変異体は通常個体と比べて約半数の花粉数しか生産しないことがわかりました。この遺伝子の変異体にさらに正常な遺伝子を導入すると花粉数が正常に回復したことから、この遺伝子を、花粉数を制御する遺伝子REDUCED POLLEN NUMBER1(RDP1)と名づけました。
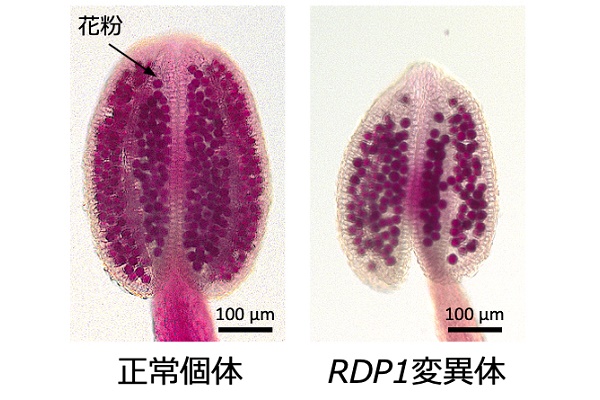
RDP1の配列によって花粉数のみを変え、生育メリットは保持
それでは、シロイヌナズナはどのようにして花粉数を変えているのでしょうか? シロイヌナズナは自家受粉を行う植物で、花粉数が少ないことは進化上有利に働くと考えられますが、遺伝子機能が破壊されたRDP1を持つ系統は見つかりませんでした。
そこでRDP1変異体をよく観察したところ、正常個体と比較して生育が遅れることがわかりました。このことから、シロイヌナズナにとってはRDP1を持たないことで花粉が少なくなるメリットよりも、他個体との生存競争に不利になるデメリットの方が大きいことが予想されます。したがって、自然界ではRDP1を持たないシロイヌナズナの個体は存在しない可能性が考えられ、さらに、シロイヌナズナはRDP1の配列を変えることで花粉数を変えている可能性が考えられました。
さらにRDP1遺伝子の周辺領域の配列を解析したところ、花粉の生産が少ない個体のグループでは、花粉を多く生産するグループと比較して変異が少ないことがわかりました。これは選択的一掃(Selective sweep)と呼ばれる現象で、自家受粉を行うシロイヌナズナではRDP1領域において花粉が少ない配列が正の選択を受けているということを示しています。
次に花粉の多い系統由来のRDP1配列と花粉の少ない系統由来のRDP1配列によって実際に花粉数が変わるかどうかを調べるために、遺伝的背景を揃え、RDP1のタイプのみが異なる個体同士を作出し、花粉数を比較しました。
その結果、花粉の多い系統由来のRDP1配列を持つ個体がより多くの花粉を生産することがわかりました。興味深いことに、これらの個体間で生育速度や花器官サイズなどには差がないことがわかりました。シロイヌナズナは自然界において、RDP1配列が変わることで花粉数のみを変えていることがわかりました。
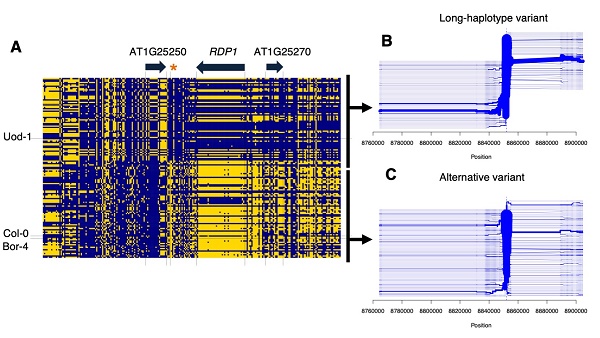
(Tsuchimatsu, T., Kakui, H. et al., より引用)
花粉数制御の遺伝子ネットワークの解明と応用に向けて
今回私たちの研究によって、初めて花粉数を制御する遺伝子RDP1を発見することに成功しました。今後は、RDP1を手掛かりに解析を進めることで花粉数を制御する新規の遺伝子を明らかにし、花粉数を決める遺伝子ネットワークを解明していきたいと考えています。
また、RDP1は基本的にどの植物にも存在する遺伝子であることがわかっており、今後さまざまな植物のRDP1について遺伝子マーカーの開発やゲノム編集を行うことで、人工授粉が必要な植物には多くの花粉を生産する個体の選抜に利用することや、花粉症の原因となる植物に関しては少花粉の個体を作出するなど植物種や分野を超えた応用が期待されます。
参考文献
- *Tsuchimatsu, T., *Kakui, H., Yamazaki, M., Marona, C., Tsutsui, H., Hedhly, A., Meng, D., Sato, Y., Städler, T., Grossniklaus, U., Kanaoka, M.M., Lenhard, M., Nordborg M., Shimizu, K.K. (2020) Adaptive reduction of male gamete number in the selfing plant Arabidopsis thaliana. Nature Communications 11,2885. *第一著者https://doi.org/10.1038/s41467-020-16679-7
- 土松隆志, 清水健太郎 (2010) シロイヌナズナ属における自家和合性進化の遺伝的背景を探る. BSJ-Review 1, 90-95.
https://bsj.or.jp/jpn/general/bsj-review/BSJreview2010B7.pdf - Kakui, H., Yamazaki, M., Shimizu, K.K. (2020) Pollen grain counting using a cell counter, Pollen and Pollen Tube Biology, 1-11, Springer US https://www.springer.com/gp/book/9781071606711
この記事を書いた人

-
角井 宏行(写真左上)
新潟大学自然科学研究科 特任助教。
2009年 千葉大学自然科学研究科にて博士課程修了。日本学術振興会特別研究員(DC2, PD, 海外)、チューリッヒ大学博士研究員、横浜市立大学・木原生物学研究所・特任助教を経て、2019年より現職。現在はスギを研究材料に無花粉スギの原因遺伝子の解析などを行っている。
土松 隆志(写真右上)
東京大学大学院理学系研究科 准教授。
2010年 東京大学大学院総合文化研究科博士課程修了。チューリッヒ大学博士研究員、グレゴール・メンデル研究所博士研究員、千葉大学大学院理学研究院准教授等を経て2020年より現職。さまざまな植物、藻類を用いて、ゲノムデータから適応進化の機構を解明する研究を進めている。
山崎 美紗子(写真左下)
チューリッヒ大学、進化生態ゲノミクス研究室所属。
2008年 京都大学理学研究科博士課程修了。ローザンヌ大学にて博士研究員を経験後、2012年より現職。日本にはまだあまりないラボマネージャーという立場から、研究が円滑に進むよう、さまざまな面からのサポートに取り組んでいる。
清水 健太郎(写真右下)
チューリッヒ大学進化生物学・環境学研究所・教授、横浜市立大学木原生物学研究所・客員教授。
2002年 京都大学理学研究科博士課程修了。アメリカ・ノースカロライナ州立大学研究員を経て、2006年 チューリッヒ大学に着任。フィールドの進化・生態学と実験室の分子遺伝学を融合して、野外環境での植物の生き様を研究してきた。