細胞性粘菌がつくり出す密度波状の集団運動 – 原動力は細胞の”追っかけっこ”だった!
細胞の集団運動はなぜ起こるか
生命の最小単位である細胞は、しばしば集団化しながら運動することがあります。特に真核細胞の集団運動は、形態形成や創傷治癒などのプロセスで重要な役割を果たしており、生命科学の広い分野で研究されています。細胞の集団運動は一般的に、一部の細胞が分泌する化学物質への「走化性」により細胞集団が誘導されたり、複数の細胞がある分子を介して強固に接着しながら一緒に運動したりすることで実現されます。このように、細胞の集団運動では、細胞集団内で起こる高度な相互作用が必要と考えられてきました。
一方、近年の物理学の研究において、「隣の個体と運動方向をそろえる」といった単純な相互作用が集団運動を可能にするということが、明らかになってきています。実際に、動物、細菌、生体高分子などのさまざまな種類の集団において、単純な相互作用による集団運動が観察されてきており、これらの集団においては、観察される集団運動と集団内の個体の挙動の関係性が明らかになっています。たとえば、分子モーターによって駆動される生体高分子集団は、個々の生体高分子同士の衝突という単純相互作用により、結果としてお互いが整列し、集団運動が引き起こされます。
しかしながら、真核細胞に目を向けてみると、このような単純相互作用による集団運動に関しては、ほとんど研究がなされていません。真核細胞集団における、走化性や強固な細胞接着などを伴わない集団運動、またその原因となる相互作用の特定は、多細胞生物内で起こる複雑な集団運動を理解する新たな鍵となる可能性があります。
走化性を示さない細胞性粘菌の集団運動
細胞性粘菌(Dictyostelium discoideum)は、細胞集団運動のモデル生物として古くから研究されています。細胞性粘菌は、ひとつひとつの細胞が独立に運動し、周囲のバクテリアなどの栄養を取り込みながら生活しますが、周囲の栄養がなくなり飢餓状態になると、細胞が集合して多細胞体制に移行します。細胞が集合する際には集団運動が深く関わっており、その集団運動は、細胞が分泌するcAMP(環状アデノシン一リン酸)というシグナル分子への走化性によって実現します。つまり、細胞間における分子の放出と受容という高度な相互作用が集団運動を引き起こします。
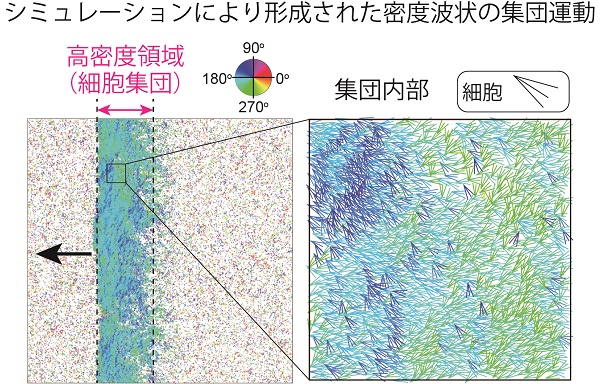
ところが、近年、走化性を示さない細胞性粘菌の変異株(KI細胞)の集団においても、集団運動が観察されることが発見されました。KI細胞を寒天培地上に均一に撒き、飢餓状態にさせても、走化性による集団運動が実現しないため多細胞体制は構築されません。その代わり、細胞密度が低い領域と高い領域とに分かれる様子が観察されることがわかっています。
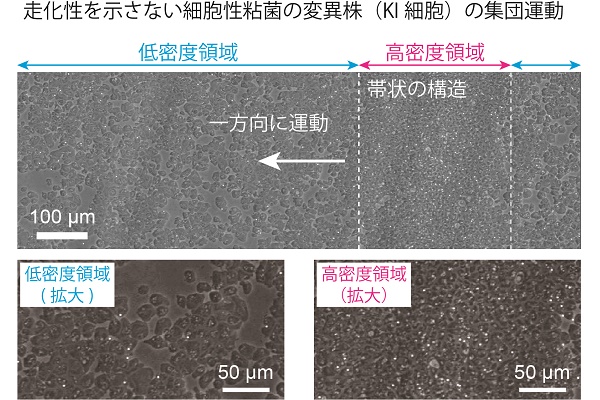
高密度領域は帯のような構造を持っており、この帯に対して垂直方向に運動するため、進行波のように見えます。これは、野生株の集団運動とは異なる、走化性に依存しない集団運動として注目されていましたが、現在までその原理はわかっていませんでした。そこで私たちは、この集団運動と細胞間相互作用を定量的に解析し、形成メカニズムを実験・数値シミュレーション両側面から明らかにすることを試みました。
KI細胞の集団運動の正体は「密度波」
私たちははじめに、この進行波様の集団運動の振る舞いを定量的に調べることにしました。まず、進行波そのものの速度は、異なるサンプル間においてもほぼ一定で、0.5μm/sでした。一方、帯状の集団内部に存在するひとつひとつのKI細胞の運動速度は、約0.4μm/sであり、集団内のKI細胞ひとつひとつの速さが、KI細胞集団が形成する進行波そのものよりも遅いことがわかりました。また、1細胞を追跡した運動軌跡からは、はじめ集団の外にいた細胞は、あるとき、集団の内部に移動して進行波として振る舞った後、再び集団の外に戻っていることが明らかになりました。
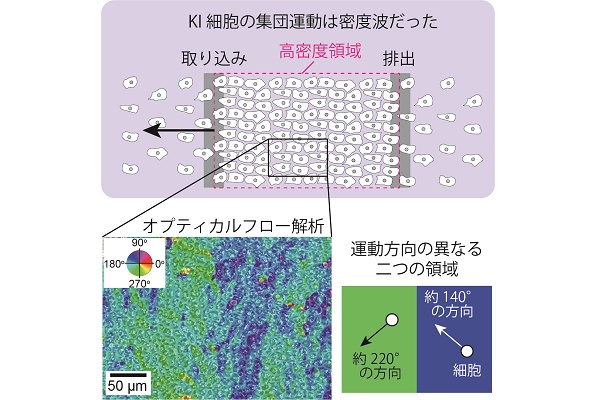
これらの観察から、KI細胞の進行波様の集団運動は、同じ細胞が集団となって移動しているのではなく、細胞の高密度領域が伝搬する「密度波」であることがわかりました。細胞は集団の先頭で集団に加わり、集団の進行とともに集団の後方へと移動し、そして後方から集団をはずれ集団の外へ戻っていくという動的な過程が繰り返し起こることで、集団は個々の細胞の運動速度よりも速い速度で移動していくことができます。
KI細胞は集団の外に存在するときは、それぞれがバラバラの方向に動いているように見える一方で、集団内にいるあいだは波の進む方向に動いているように見えます。そこで、細胞極性(運動方向)がどれだけそろっているかを定量的に解析するため、秩序変数(order parameter)と呼ばれる値を、集団内外に対してそれぞれ算出しました。すると、秩序変数は集団内においてのみ高い値を示しました。つまり、集団内の細胞極性は周囲とそろっており、集団内の細胞は比較的同じ方向に動いていることがわかりました。
さらに、集団運動をより詳しく調べるために、オプティカルフローと呼ばれる解析法を用いて解析を行った結果、集団内に運動方向の異なる2つの領域が存在しました。波の進行方向を180°とした場合、左斜め下(約220°方向)に運動する集団と左斜め上(約140°方向)に運動する集団とが、ストライプのように交互に存在した構造が見られました。
集団運動は接触追随運動によって起こされる
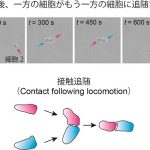
冒頭でも示したように、一般的に、集団運動を実現するためには細胞間における何らかの相互作用が必要であると考えられます。そこで、細胞が1対1で衝突した後、どのように相互作用するかを観察しました。この結果、一方の細胞がもう一方の細胞に追随するような現象が見られました。私たちはこの現象を、接触追随(contact following locomotion:CFL)と名づけました。
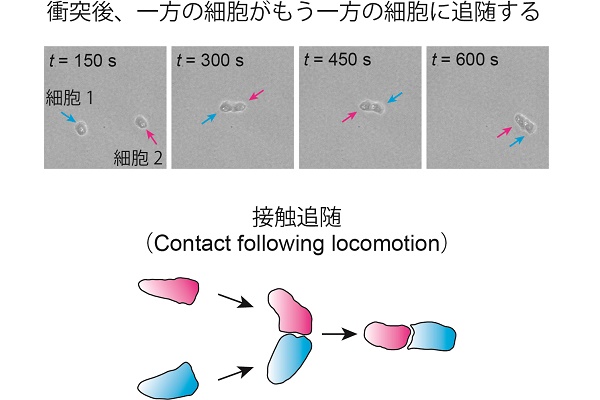
実は、CFLのような「細胞が細胞の後を追っていく」といった現象はKI細胞に限らず、野生株の細胞性粘菌においても観察されています。また、TgrB1というタンパク質がCFLを担うタンパク質のひとつであることも知られています。実際に、私たちがtgrb1遺伝子を発現できないKI細胞変異株を用いて解析を行ったところ、密度波状の集団運動は形成されないことを確認しました。また、この変異株を用いて、細胞が1対1で衝突した後の細胞間相互作用を調べたところ、CFLの形成はほとんど認められませんでした。したがって、密度波状の集団運動は細胞間におけるCFLに起因するものであると結論づけられました。
さらに、私たちはCFLが密度波状の集団運動を引き起こすかどうかを、エージェントベースの数理モデルを構築して確かめました。この数理モデルにおいて、各エージェントは位置と極性の変数で記述され、各エージェントはそれぞれの極性の方向に運動します。CFLの効果は、エージェントが隣接するエージェントの後方にあるときに、極性がその方向に向く作用として導入しました。
この数理モデルを用いてシミュレーションを行ったところ、ある条件でCFLの強さに依存して密度波状の集団運動が形成されました。密度波の進行速度と、集団運動を形成しているエージェントの運動速度を比較したところ、前者の方が速いという実験と同じ結果が得られました。また、集団内に観察された運動方向の異なる領域も、細胞性粘菌での実験と同様に観察されました。これらの結果から、CFLによって密度波状の集団運動が誘導されることが確かめられました。
密度波状集団運動の意義解明に向けて
私たちの研究によって、KI細胞が示す密度波状の集団運動は、一方の細胞がもう一方の細胞に追随することに起因すること、さらに、運動している細胞集団内に運動方向の異なる2つの領域がストライプのように交互に存在していること、そして、この構造はCFLによって引き起こされるパターン形成であると考えられることが明らかになりました。しかしながら、何のために、どのようにして、この構造ができ上がるのか? という疑問への回答は今後の課題として残っています。
野生株細胞性粘菌は、多細胞体制に移行するために集合する際、走化性に基づいた集団運動を示しますが、集合後も多細胞体内部において細胞が集団運動していることが知られています。ごく最近、多細胞体内部における集団運動には、走化性ではなく、他のメカニズムが重要な役割を果たしていることが示唆されています。私たちは、CFLによって誘導される集団運動が、このような細胞性粘菌の形態形成に寄与していると考えています。
またCFLは、細胞性粘菌に限らず、哺乳類細胞においても報告されています。したがって、本研究で報告した密度波状の集団運動は、他のシステムでも観察される可能性があり、今後生命科学の分野全体の理解に広く貢献できると期待しています。
Masayuki Hayakawa, Tetsuya Hiraiwa, Yuko Wada, Hidekazu Kuwayama, and Tatsuo Shibata, “Polar pattern formation induced by contact following locomotion in a multicellular system”,eLife, 9: e53609. DOI: 10.7554/eLife.53609
この記事を書いた人

-
早川 雅之(画像左)
理化学研究所フィジカルバイオロジー研究チーム・研究員。
2017年東京工業大学大学院総合理工学研究科知能システム科学専攻にて博士(理学)を取得後、日本学術振興会特別研究員(PD)を経て、2020年から現職。
柴田 達夫(画像右)
理化学研究所 生命機能科学研究センター フィジカルバイオロジー研究チーム チームリーダー。
東京大学大学院総合文化研究科博士課程修了、博士(学術)、京都大学数理解析研究所研究員、ドイツ・マックスプランク研究機構フリッツハーバー研究所・フンボルト財団研究員、広島大学院理学研究科准教授、理化学研究所ユニットリーダーを経て現職。専門は細胞や発生の生物物理学や数理生物学。