精神神経疾患に関わるタンパク質「リーリン」 – 受容体を使い分けて神経細胞の”集合”と”整列”を制御する分子
神経細胞は動いて脳をつくる
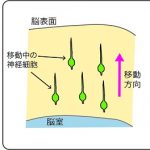
脳の機能は、多くの神経細胞が作るネットワークによって担われています。そのネットワークが正しく作られるためには、脳の形成過程において神経細胞が適切な位置に配置されることが重要です。脳が作られる際には、神経細胞は脳の深部にある脳室と呼ばれる空間に面した場所で誕生し、脳の表層に向かって移動します。移動している神経細胞は、細胞周囲の環境から多様な情報を受け取りながら、正しい場所へと配置されることが知られています。
近年、さまざまな精神神経疾患の背景に、脳の形成過程における神経細胞の配置異常などの微細な障害が存在している可能性が注目されており、神経細胞の移動と配置のメカニズムは重要な研究課題となっています。
神経細胞の挙動をコントロールする分子「リーリン」
脳の最表層に豊富に存在する細胞外糖タンパク質リーリンは、さまざまな精神神経疾患との関連が知られており、移動する神経細胞に働きかけてその挙動をコントロールする働きを持ちます。リーリンを欠損したマウスでは正常な細胞移動が行われず、またリーリンを欠損したヒト患者では脳のしわが失われる滑脳症となり、重篤な小脳形成不全も呈します。
リーリンは2種類の受容体ApoER2、VLDLRに結合して働きます。リーリンの機能は、神経細胞を特定の場所に移動させ集める「集合」と、適切な場所に留置する「停止」の両方の働きをすることがわかってきましたが、これらの受容体における働きの違いについては解明が遅れていました。
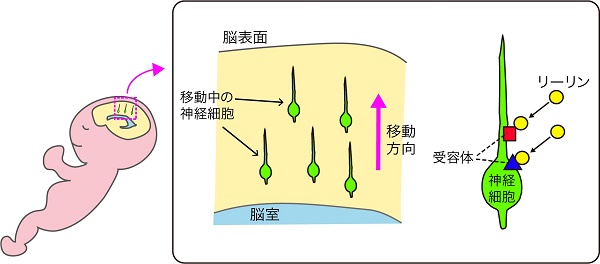
受容体を使い分けることで、同じ分子で異なる働きを実現している
私たちの研究グループは以前、形成途中のマウスの脳に、リーリンを人為的に本来とは異なる場所で(異所的に)発現させると、神経細胞が異所的に発現したリーリンの周囲に集まって細胞凝集塊を作ることを発見しました。その際、神経細胞の細胞体はリーリンが濃縮して存在する中心部分には進入せず、その周囲に規則正しく整列して凝集塊を作ることも明らかにしました。
また、私たちの先行研究において、リーリンの受容体のうちApoER2を欠損したマウスでリーリンを異所発現させると、神経細胞はリーリンに反応せず集合しないことを見出しました。さらに今回の研究では、もうひとつの受容体であるVLDLRを欠損したマウスの解析によって、神経細胞は異所発現されたリーリンに反応して集合するものの、正しい位置に停止できず不規則に並ぶことがわかりました。
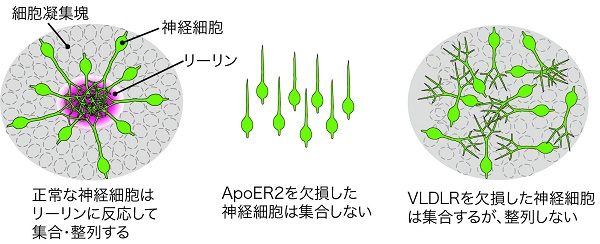
それぞれの受容体を欠損したマウスにおいて脳の発生過程を調べたところ、ApoER2を欠損したマウスでは、神経細胞が移動を開始して間もなく移動の効率が悪くなり、多くの細胞が本来の目的地である脳表層まで到達できないことがわかりました。また、脳表層まで到達できた細胞についても、一部の細胞は正常に集合・配置されないことも観察されました。一方、VLDLRを欠損したマウスの発生過程の脳では、神経細胞は脳表層まで到達しますが、本来の位置で停止せずそれを越えてしまう細胞がいることもわかりました。
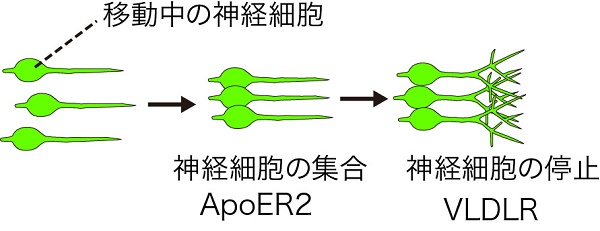
これらの結果は、リーリンは脳の表層まで移動してきた神経細胞に対して、まずApoER2を介して集合させ、次にVLDLRを介して正しい位置に留まらせて整列させることを示しており、同じ分子が2種類の受容体を使い分けることで別の働きをしていることが明らかになりました。
脳作りの基盤メカニズム解明に向けて
神経細胞が適切に移動して脳を形作るためには、多数のステップが厳密にコントロールされることが不可欠であり、その各ステップの破綻は重篤な疾患を引き起こします。これまでの研究では、リーリンの2種類の受容体は脳の発生過程において似た機能を重複して持っていると考えられていましたが、今回の研究によって、同一の分子が神経細胞の集合と配向決定という別個の重要なステップを、2種類の受容体を使い分けることによってコントロールしていることが明らかになりました。
今後、リーリンによる受容体の使い分け制御の仕組みを解明することで、脳作りの基盤メカニズムの理解につながると期待されます。さらに、神経細胞の配置異常が背景に存在するとして注目されている滑脳症をはじめとする各種精神神経疾患の新たな病態の解明や、治療法の開発にも貢献できる可能性があると考えられます。
参考文献
- Hirota, Y., Kubo, K., Fujino, T., Yamamoto,T., Nakajima, K. ApoER2 controls not only neuronal migration in the intermediate zone, but also termination of migration in the developing cerebral cortex. Cerebral Cortex. 28:223-235. 2018.
- Hirota, Y. and Nakajima, K. VLDLR is not essential for Reelin-induced neuronal aggregation but suppresses neuronal invasion into the marginal Development. 147: dev189936. 2020.
この記事を書いた人

-
廣田 ゆき(画像右)
慶應義塾大学医学部解剖学教室・専任講師。
2003年大阪大学大学院医学系研究科博士課程修了。慶應義塾大学研究員、学振特別研究員、名古屋市立大学助教、慶應義塾大学助教を経て、2016年より現職。脳の発生・発達・病態を制御するメカニズムの研究を行っています。
仲嶋 一範(画像左)
慶應義塾大学医学部解剖学教室・教授。
1994年大阪大学大学院医学系研究科博士課程修了。学振特別研究員、理化学研究所ライフサイエンス筑波研究センター研究員を経て、1998年東京慈恵会医科大学DNA医学研究所部門長・講師として独立。同准教授を経て2002年より現職。