“葉緑体ドロボウ”は進化途中の現象か!? – 盗葉緑体性渦鞭毛藻「ヌスットディニウム」の細胞内共生の仕組み
細胞内共生は多様な藻類を生み出す原動力
陸上の植物は、細胞内の葉緑体によって光エネルギーを利用して空気中の二酸化炭素を固定し、有機物を作る光合成を行います。葉緑体は10億年以上前、植物の祖先が自分の細胞内に共生させたシアノバクテリアに由来しています。
細胞内共生の過程には未だに多くの謎が残されていますが、葉緑体を持っていなかった祖先の細胞内に、シアノバクテリア様の細胞がそのまま入り込んで(あるいは食べられて)、やがて葉緑体になったと説明されています。
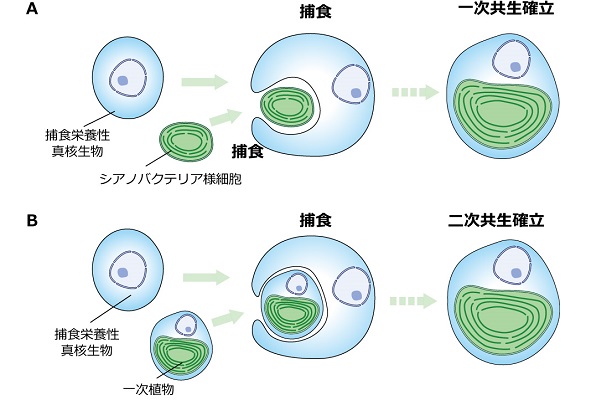
(A)一次共生の進化。葉緑体は、捕食栄養性だった祖先真核生物がシアノバクテリア様細胞を取り込んで、細胞内に住まわせることに由来するオルガネラである。
(B)二次共生の進化。「藻類」の葉緑体の中には、捕食栄養性だった祖先の真核生物がシアノバクテリアを共生させた祖先に由来する一次植物を取り込んで、細胞内に住まわせることに由来するものもある。
水圏には「藻類」と呼ばれる光合成をする生物が豊富に生息しています。池や湖、海の水を掬って顕微鏡で観察すると、多種多様な形態の「藻類」を目にすることができます。海水浴や日常の食卓で見かける海藻も「藻類」と呼ばれます。それらは陸上植物と同様に、光エネルギーと二酸化炭素、無機栄養塩などから自らを形作る有機物を産生し、生育することができます。
これらのプロセスは葉緑体で行われる光合成によるものですが、「藻類」の葉緑体はシアノバクテリアを共生させた祖先から直接受け継いだものとは限りません。たとえば、ミドリムシ類(ユーグレナ類)、褐藻類・珪藻類(不等毛藻類)は陸上植物の系統とはそれぞれまったく異なる系統に属する「藻類」です。
このような多様性は、「藻類」がそれぞれの系統で独立に誕生するイベントが何回も起こったことに由来しています。「藻類などを食べて生きていた生物が、藻類を自分の細胞に住まわせることに成功し、自らも藻類になる」という劇的な進化が、藻類の多様性を生み出す原動力になっているのです。
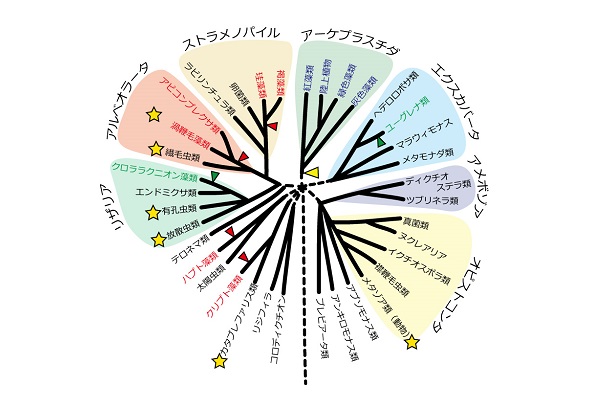
青色文字の系統群は、シアノバクテリアを共生させた一次共生に由来する葉緑体を持つ系統群である。赤色と緑色の文字の系統群は、それぞれ紅藻と緑藻を共生させた二次共生に由来する葉緑体を持つ系統群である。系統樹の枝の黄色、赤色、緑色の矢尻は、それぞれシアノバクテリア、紅藻、緑藻との細胞内共生を果たしたタイミングを示す。
藻類を食べて新たな藻類が作り出される! – 真核生物の藻類化の仕組み
では、ミドリムシ類や不等毛藻類はどのようにして葉緑体を手にしたのでしょうか。シアノバクテリアとの細胞内共生は一次共生と呼ばれており、陸上植物を含む緑色植物、紅藻類等は一次共生によって葉緑体を獲得した祖先から進化しました。
一方、一次共生で葉緑体を獲得した真核藻類(一次植物)と真核生物宿主の共生は二次共生と呼ばれます。ミドリムシ類は緑藻との二次共生を経て自らも藻類になったグループです。不等毛藻類の祖先も紅藻との二次共生により葉緑体を手にしました(葉緑体の由来、共生の回数については諸説あり)。このように、葉緑体を獲得する進化は、原核生物と真核生物の共生だけでなく、真核生物と真核生物の共生でも起こり得ることなのです。
クリプト藻類は不等毛藻類とは全く異なる系統の藻類です。クリプト藻は不等毛藻類とは独立に紅藻と細胞内共生を果たして藻類化したグループであり、真核生物を共生させたという確固たる証拠が残されています。
クリプト藻の葉緑体のそばには、真核生物の核にそっくりな構造があります。これはヌクレオモルフと呼ばれており、約500遺伝子がコードされた染色体が維持されています。500という遺伝子の数はひとつの生物を維持するのにはあまりにも少ない数ですが、共生した「元」藻類を維持する残りの遺伝子は、クリプト藻本体の核に移行し、それらを使って宿主細胞が共生藻の維持・管理をしていると説明されています。
クリプト藻内の共生藻はもはやクリプト藻の細胞の一部にまで統合され、自律性を失ってしまっていますが、このヌクレオモルフの存在は、真核生物を取り込む細胞内共生が確かに起こったという証拠なのです。
「葉緑体ドロボウ」は葉緑体獲得の途中段階なのか?
真核生物内で「藻類化」が多数回起こったということは通説になっています。それらの細胞内共生は、太古の昔に2つの生物だったものが、すでに数億年前にひとつの生物として完成したものです。現在の「藻類」はそういった生物の子孫であり、宿主細胞と葉緑体は強固に結びついている“永続的な”関係性にまで進化しているため、細胞内共生の途中で、何がどのような順序で起こったのかということを推測するのは難しいと考えられます。
このようななか、永続的に維持される共生藻が確立されるまでの過程の謎に対して光明を投ずるかもしれない現象が、「盗葉緑体」現象です。盗葉緑体現象は、もともと葉緑体を持っていない生物が捕食によって藻類を取り込んで、その葉緑体を一時的に維持する現象のことを指し、真核生物の多くの系統に確認されています。
宿主は葉緑体を維持しているあいだ、盗んだ葉緑体で光合成を行いますが、やがては葉緑体を消化してしまいます。このような藻類を食べてはその葉緑体を一時的に細胞内共生させるというサイクルは、現在の「藻類」が太古の昔に経験した、捕食して栄養を得ていた生物が真の葉緑体を獲得しようとしている中間の段階なのではないかと考えられています。
私はこれまで、「葉緑体を捕食して生きていた生物がどのようにして葉緑体を獲得するのか」という点に興味を持ち、盗葉緑体現象を示す渦鞭毛藻類、その名もヌスットディニウム属の渦鞭毛藻類を使って研究してきました。これらの渦鞭毛藻類はクリプト藻を取り込んでその葉緑体を盗む生物で、名前の「ヌスット」は文字どおり日本語の「盗人」から来ています。ヌスットディニウム・アエルギノーサムという種は細胞全体に行き渡るほどの大きさの葉緑体を持っています。
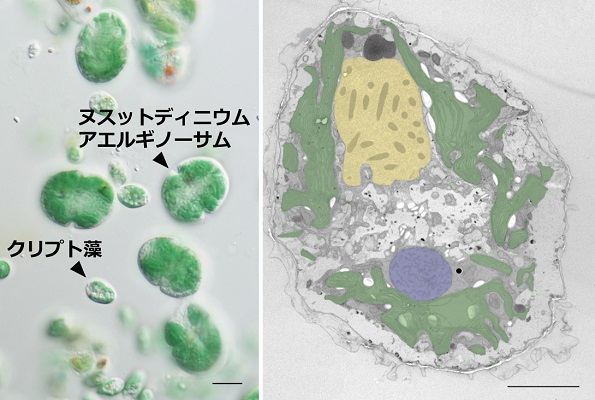
(左)ヌスットディニウム・アエルギノーサムとその餌のクリプト藻の光学顕微鏡写真。スケールバーは10μmを示す。
(右)ヌスットディニウム・アエルギノーサムの透過型電子顕微鏡写真。強調部位は、拡大されたクリプト藻の盗葉緑体(緑色)、クリプト藻核(青色)、渦鞭毛藻核(黄色)を示す。スケールバーは5μmを示す。
私が研究に着手する以前は、この葉緑体がクリプト藻に由来するものであるということはわかっていましたが、取り込まれる前のクリプト藻の葉緑体よりも遥かにサイズが大きく、このような形態になる理由は不明でした。私は、ヌスットディニウム属の盗葉緑体現象が、捕食の段階から葉緑体を獲得する段階に至るまでにどのような現象が起こったかを解明するための良いモデルになると考え、以下のような研究を行いました。
ヌスットディニウムの葉緑体サイズの維持に盗んだ核が重要
私が研究生活を始めた当時、ヌスットディニウム属渦鞭毛藻類の培養株がなかったため、渦鞭毛藻の採集と培養株の作成に着手しました。堀口健雄教授の研究室(北海道大学理学院)の所属時に培養株の作成に成功し、細胞内の盗葉緑体の変化を透過型電子顕微鏡観察で明らかにしようと試みました。
葉緑体を完全に消化させた渦鞭毛藻細胞を作り、1細胞のクリプト藻を食べさせて、その後の変化を観察しました。クリプト藻はまるごと渦鞭毛藻に取り込まれ、葉緑体と核などが細胞内に維持されます。
その後、クリプト藻の葉緑体は、渦鞭毛藻細胞の全体に行き渡るほどの大きさになり、元のサイズの20倍以上に拡大されることが明らかとなりました。さらに驚くべきことに、拡大された葉緑体は渦鞭毛藻の細胞分裂と同調して分裂し、両方の娘細胞に受け継がれることも明らかとなりました。しかし、クリプト藻の核は渦鞭毛藻細胞と同調して分裂せず、片方の細胞のみに受け継がれていくこともわかりました。
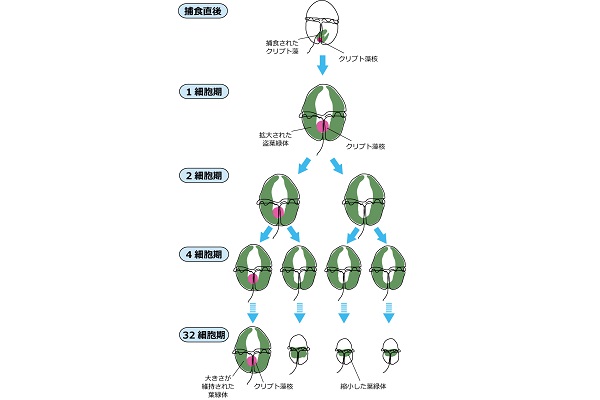
クリプト藻は渦鞭毛藻にまるごと取り込まれ、葉緑体とクリプト藻核等が維持される。クリプト藻の葉緑体は72時間後には細胞全体に拡大され、120時間ほどで渦鞭毛藻細胞が細胞分裂を開始する。盗葉緑体は渦鞭毛藻細胞と同調分裂し、両方の娘細胞に受け継がれるが、クリプト藻核は分裂せずに片方にのみ受け継がれる。クリプト藻核を失った細胞は葉緑体が縮退し、クリプト藻核を維持する細胞のみが大きな盗葉緑体を持つ。
さらに細胞分裂を繰り返した渦鞭毛藻細胞の盗葉緑体とクリプト藻核を追跡すると、クリプト藻核を受け継いだただひとつの細胞が常に細胞全体に渡るサイズの葉緑体を維持しているのに対し、クリプト藻核を失った細胞は葉緑体の拡大を伴わない分裂を繰り返すために、葉緑体が縮退していくことがわかりました。このことから、盗葉緑体の維持・拡大にはクリプト藻核が必要であり、クリプト藻核の遺伝子が使われている可能性があることが示唆されました。
宿主細胞内では劇的な共生藻リモデリングが起こっている!
私が国立遺伝学研究所の宮城島進也教授の研究室(共生細胞進化研究室)にポスドクとして異動してから、盗葉緑体現象のメカニズムを明らかにすべく、次世代シーケンサーを用いたトランスクリプトーム解析や種々の培養実験に取り組んできました。トランスクリプトーム解析とは、細胞内の転写物(mRNA)を網羅的に配列取得することで細胞内の状態を推測する解析です。この解析によって、細胞内のクリプト藻核の働きや、渦鞭毛藻に取り込まれた前後での変化を明らかにしようと試みました。
その結果、渦鞭毛藻に取り込まれたクリプト藻核は、形態観察で示唆されていたように、転写活性を維持しており、取り込まれる前よりも代謝・翻訳・DNA合成に関する遺伝子群の発現が上昇することが明らかとなりました。このことから、渦鞭毛藻内のクリプト藻は、光合成や光合成産物の代謝、葉緑体の製造に特化した“マシーン”のように変化していると予想されます。
さらに、渦鞭毛藻に取り込まれる前には明暗の切り替えに即座に反応して転写を変化させていたクリプト藻核が、取り込まれた後は明暗の切り替えに応答した遺伝子発現の変化がなくなることもわかりました。渦鞭毛藻内のクリプト藻は取り込まれる前に持っていた細胞外皮構造や眼点、鞭毛等を失うので、外界の刺激に対する応答ができなくなっていると考えられます。
また、クリプト藻核が渦鞭毛藻細胞内に維持されているあいだ、核のDNA量が増えていくことも明らかになりました。前述のとおり、クリプト藻核は渦鞭毛藻細胞内で分裂しなくなります。その状況でDNA複製が起こることは、すなわちクリプト藻核が多倍体になっていることを意味しています。このように、トランスクリプトーム解析からも渦鞭毛藻に取り込まれたクリプト藻には劇的な変化が起こっていることが明らかとなりました。
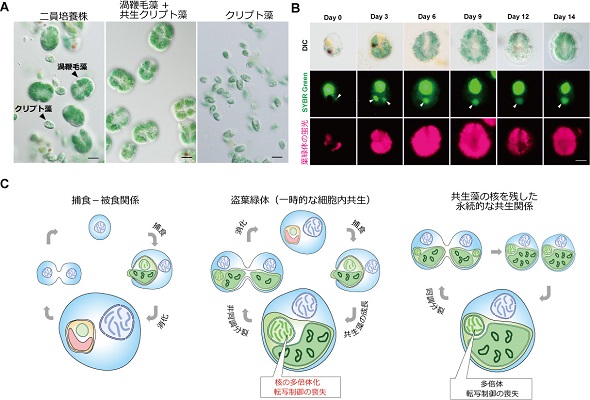
(A)渦鞭毛藻とクリプト藻の二員培養(左)、クリプト藻を取り込んで維持している渦鞭毛藻(中央)、取り込まれる前のクリプト藻(右)の写真。スケールバーは10μm。
(B)渦鞭毛藻に取り込まれた後のクリプト藻核(矢頭:緑の蛍光)と葉緑体(赤の蛍光)の変化。渦鞭毛藻に取り込まれたクリプト藻核と葉緑体は徐々に拡大する。スケールバーは10μm。
(C)本研究の結果から示唆される、捕食-被食関係から永続的な共生関係への進化過程。(左)捕食-被食関係では、捕食者は被食者をすぐに消化する。(中央)ヌスットディニウム・アエルギノーサムに見られる盗葉緑体現象。取り込まれた藻類核の多倍体化と転写制御能の喪失が起こる。(右)葉緑体とその起源となった細胞の縮退核を持つ一部の藻類でみられる永続的な共生関係の維持。葉緑体と多倍体化した縮退核を自身の細胞分裂に同調させており、環境変動に応じた転写制御がなくなっていることが知られている。
しかし、細胞内のクリプト藻は単に渦鞭毛藻の奴隷のように扱われている訳ではありません。渦鞭毛藻を強光下で培養すると、クリプト藻核を失った細胞は強光下で速やかに死滅することから、クリプト藻核の遺伝子群は光合成酸化ストレスへの対応の役割も果たしていることが示唆されました。
このように、クリプト藻核は光合成酸化ストレスを生じる強光条件下で宿主渦鞭毛藻の生存を支えていることが明らかとなり、渦鞭毛藻は盗葉緑体だけではなく、クリプト藻核にも強く依存していることが示唆されます。ヌスットディニウム・アエルギノーサムにとっては、自分自身をひとつの生物として維持するために、クリプト藻がなくてはならない存在となっていると考えられます。
興味深いことに、ヌスットディニウムの盗葉緑体現象における、取り込まれた藻類核の多倍体化と転写制御の喪失は、前述のヌクレオモルフ、そしてあらゆる「藻類」の葉緑体、その他種々の共生現象にも共通しています。つまり、これらの現象は、永続的な共生関係を確立する前から起こり得て、真の葉緑体を持つ生物群も太古の昔には盗葉緑体性生物であった可能性が示唆されます。
参考文献
- Keeling PJ. The number, speed, and impact of plastid endosymbioses in eukaryotic evolution. Annu Rev Plant Biol 2013; 64: 583–607.
- Onuma R, Horiguchi T. Kleptochloroplast enlargement, karyoklepty and the distribution of the cryptomonad nucleus in Nusuttodinium (= Gymnodinium) aeruginosum (Dinophyceae). Protist 2015; 166: 177–195.
- Onuma R, Hirooka S, Kanesaki Y, Fujiwara T, Yoshikawa H, Miyagishima S. Changes in the transcriptome, ploidy, and optimal light intensity of a cryptomonad upon integration into a kleptoplastic dinoflagellate. ISME J 2020; https://doi.org/10.1038/s41396-020-0693-4
この記事を書いた人

-
日本学術振興会特別研究員PD(受入は国立遺伝学研究所 共生細胞進化研究室)。
2010年、山形大学理学部生物学科卒業。2015年、北海道大学院理学院自然史科学専攻博士後期課程終了。博士(理学)。2015年、国立遺伝学研究所博士研究員を経て2018年より現職。