種分化は大きな島で起きやすい – 琉球列島で5回独立に平行進化した「キバラヨシノボリ」から多様化の地理的条件を探る
種分化の起きやすさと生態系サイズの関係とは?
地球上にはさまざまな生物種が存在していますが、その多様性の度合は場所ごとに大きく異なります。たとえば赤道直下の熱帯雨林では多種多様な生物がひしめき合ういっぽうで、北極や南極にはとても限られた種しか分布していません。このような種多様性のばらつきは、他にも標高間や、大陸と島のあいだなど、さまざまな地域間で見られます。
この種多様性の地理的なばらつきを説明する仮説のひとつとして、1種が2種以上に分かれる現象である「種分化」の起こりやすさの違いを原因と考えるものがあります。すなわち、種多様性の高い地域は種分化が起こりやすく、多様性の低い地域では起こりにくいためだという仮説です。
では種分化の起こりやすさに、場所ごとの違いを与える要因はいったい何でしょうか? これまでにさまざまな潜在的な要因が挙げられてきましたが、なかでも説明力の高いもののひとつとして、生息場所の生態系サイズが知られています。ここでいう生態系サイズとは、島の面積、湖の体積や深さ、などです。生態系サイズが大きい場所では種分化が起こる確率が高く、それが小さい場所では種分化が起こりにくい、というパターンがいくつかの生物群で知られています。
ではさらに踏み込んで、この種分化の確率と生態系サイズの正の相関が生まれるメカニズムは何でしょうか? 仮説はいろいろ挙げられていますが、明確に特定されたことはありません。種分化は、個々の分岐イベントごとに異なるプロセスを経て進行します。たとえば、ある場合には地理的な隔離障壁ができることで、また別の場合には地理的に移動が可能であるにも関わらず、異なる環境に適応した結果として、種分化が進行します。
これまでの研究では特定の生物群に属する、異なるプロセスを経た種分化をすべてまとめたうえで相関が調べられるのが普通でした。個々の種分化のどのようなプロセスが生態系サイズと相関して起こりやすかったのかを検討するためには、対象とする生物群に含まれるすべての種分化の過程を検討する必要があり、実際は非常に困難です。種分化確率と生態系サイズの相関メカニズムの解明は、この点がネックとなっていました。
淡水性のキバラヨシノボリは、回遊性のクロヨシノボリから平行種分化したか?
ここで私の相棒であるヨシノボリを紹介します。ヨシノボリの仲間はハゼ科の淡水魚で、日本全国の淡水域に広く分布しています。身近な淡水魚ですので、ご存知の方も多いのではないでしょうか。ヨシノボリ属は日本で17種以上が知られ、多くの種は川で孵化した仔魚が直ちに海へ下り、数か月を海で過ごした後に川に帰ってくる両側回遊性の生活史を送ります。いっぽうで、一生を河川で過ごす淡水性の種も少なからず知られています。
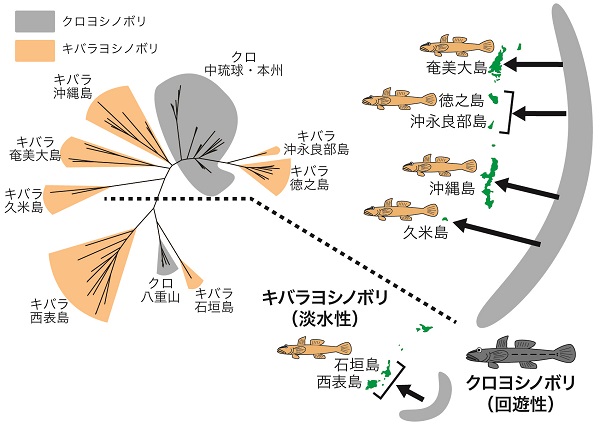
なかでも興味深いのが、琉球列島に分布する淡水性種の「キバラヨシノボリ」と回遊性種の「クロヨシノボリ」です(以降それぞれキバラとクロと呼びます)。両種は最近縁で、クロからキバラが進化したと考えられています。不思議なことに、淡水性のキバラは海を移動できないにも関わらず、琉球列島の複数の島によく似た見た目の種が分布しています。その説明として、キバラがクロから地点ごとに何度も進化した可能性が指摘されていました。
このように、場所は異なるが類似した環境で独立に適応進化が進行した結果、類似した形質が進化する現象は「平行進化」と呼ばれます。それが類似した種の進化であった場合には、特に「平行種分化」と呼ばれます。キバラが平行種分化した種であるという仮説の大元となるアイデアは、1960年代にすでに別のヨシノボリ類で提示されており、その後にも何度か言及されていました。しかし分布範囲を網羅し、厳密な遺伝解析をもとにこの仮説が検証された例はありませんでした。

ここで私は、キバラとクロからなる系であれば、種分化確率と生態系サイズの相関の理由を詳細に議論できる可能性に気付きました。キバラが平行種分化していた場合には、進化の方向性は「回遊性→淡水性」でそれぞれ共通することになります。そのため、回遊性種から淡水性種が分化する共通のプロセスのうち、一体どこが面積と相関して起こりやすかったかを比較的容易に検討できるはずです。そこで私は、キバラは本当に平行進化した存在なのか、もしそうならば、キバラの進化しやすさは生態系サイズで説明されるのかどうか、を検証することにしました。
島行脚と難航した解析
キバラヨシノボリは琉球列島の7つの島に分布しています。そこで当時大学院生だった私は、民間助成金やティーチングアシスタントで得た資金でそれらの島々を巡り、キバラを集めるところから始めました。
キバラは河川上流の渓流部に分布し、その多くは滝の上流側にあります。車で下流部に着いた後、しばらく沢登りが必要な場合もありました。さらにキバラの生息地はハブの仲間がよく現れるため、周囲の警戒に神経を磨り減らさねばなりませんでした。しかしいっぽうで、キバラの生息地はいずれも人工物のない、極めて自然状態の高い場所であったため、毎度心が洗われました。幸いなことに現地に詳しい方々の協力を得ることができたため、効率良く調査を進めることができました。最終的に52地点から得た1400個体以上について、遺伝的多型のデータを取得することができました。
次に解析が問題となりました。平行種分化を遺伝解析データから示すのは、実は非常にやっかいな問題を含んでいます。というのも、種分化は時として交雑を伴いながら進行します。この交雑を考慮しないと、実際には平行種分化していなくてもそのように検出してしまう可能性が指摘されていました。そこで「近似ベイズ計算法」と呼ばれる解析を採用し、交雑を考慮したシミュレーションを行うことでこの問題に対処することにしました。しかし解析用ソフトウェアが極めて難解だったため、ソフトを触り始めてから問題なく解析が行えるようになるまで、結局2年を要しました。
膨大な回数のシミュレーションを行った結果、キバラヨシノボリは琉球列島の7つの島のうち、5つの島で独立に種分化したことが支持されました。隣接した島間で起源を共有する場合もありましたが、これはおそらく氷河期に海水面が下がり、島が陸続きになっていたときに分布を広げたためと考えられました。
そしてどのような島でキバラが種分化しやすかったかを解析したところ、種分化は面積の大きい島で起きていたことが明らかになりました。キバラの場合もこれまでの例と同様に、種分化の確率と生態系サイズが強い正の相関を示したのです。
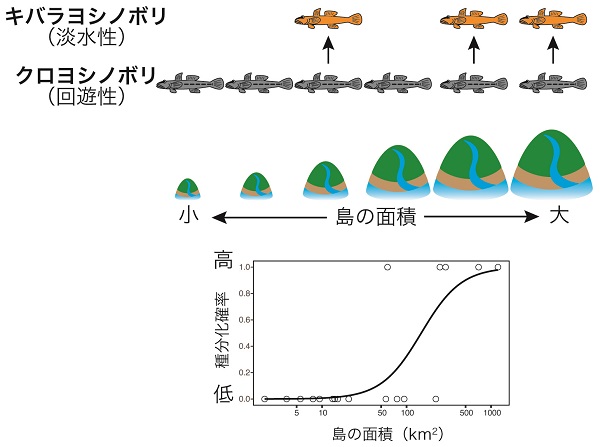
大きな島で種分化が起きやすい理由とは?
なぜキバラは大きな島で種分化しやすかったのでしょうか? 私たちは3つのメカニズムを考えています。
ひとつは島が大きいほど淡水適応と種分化を促進する選択圧が強くかかった、という仮説です。クロは川で産卵しますが、仔魚が海で小型の餌を食べて育つ必要があるため、孵化直後に流下します。クロの孵化仔魚は、海に比べ大型の餌しかない河川では餌を得られず、河川滞在期間が長いと飢えで減耗していくことが知られています。つまりクロが効率良く子を残すには下流寄りで産卵する必要があります。そのため、もし島サイズが大きく河川が長い場合には、クロが利用できない淡水域が河川上流域に生まれることになります。
このとき淡水域で生活史を完結できるような形質が現れれば、その形質が選択されることで河川上流部に適応した新たな集団が成立すると考えられます。実際にキバラの孵化仔魚はクロに比べ大きいことが知られ、これは河川の大型の餌を得るための適応だと考えられています。そして最終的に、異なる環境に適応した集団間で交配が起こらなくなることで、別種となったと考えられます。
2つ目は、生息場所が大きいほど個体数が増加するため絶滅のリスクが低下する、という仮説です。現在キバラが分布しない島であっても、過去にはキバラが分布し、島が小さいためにそれが絶滅してしまった可能性があります。その結果、現在、種分化確率と面積の相関が見られるのではないかと考えられます。
3つ目に、河川が長くなるほど河川の障害物、たとえば滝などが増えることが考えられます。琉球列島の河川には滝が頻繁に見られ、これが魚止めとなることでクロやその他の魚類とキバラの生息地を分断していることがよくあります。滝の上流部では魚類の捕食者や競争者の影響をほとんど受けないため、キバラがより安定的に集団を維持できた可能性があります。
最後に
進化学では、遺伝子頻度の変化など、種分化より前の集団内進化プロセスをmicroevolution、種分化後の多様性形成パターンをmacroevolutionと呼んで区別することがあります。これまでの種分化研究では、microevolutionとmacroevolutionは別々に追究されることが多く、遺伝子頻度の変化などのmicroevolutionの過程が、最終的にmacroevolutionのスケールでの多様性パターンの形成にどのように影響するのかはよくわかっていません。
今回の研究は、キバラの「淡水適応と種分化」というmicroevolutionのプロセスが、最終的に「種分化確率と島面積の相関」というmacroevolutionのパターン形成に関わっていたことが示唆されました。もしこの問題の解決に少しでも貢献できていれば嬉しいと思っています。
今回のキバラの種分化確率と島面積の正の相関メカニズムの説明は、残念ながら仮説の域を出ていません。今後、キバラとクロの種分化が、本当に環境適応に起因するものなのかを示すことができれば、上述の種分化確率と面積の相関メカニズムの検証に繋がるのではと考えています。また、淡水性の種が繰り返し進化した遺伝的な基盤、つまり類似した各島のキバラの進化に同じ遺伝的変異が使用されたのか、それともまったく異なる変異が利用されたのか、にも興味があります。近年ますます容易になっているゲノム解読技術を利用し、この謎を明らかにしていきたいと考えています。
参考文献
- 西田睦 “大卵化へのプロセス:ヨシノボリ類”(『水生動物の卵サイズ 生活史の変異・種分化の生物学』pp. 149-170、後藤晃・井口恵一朗(共編)、海游舎、2001)
- Kisel, Y., Barraclough, T.G. (2010) Speciation has a spatial scale that depends on levels of gene flow. American Naturalist, 175, 316-334. Doi:10.1086/650369
- Yamasaki, Y.Y., Takeshima, H., Kano, Y., Oseko, N., Suzuki, T., Nishida, M., Watanabe, K. (2020) Ecosystem size predicts the probability of speciation in migratory freshwater fish. Molecular Ecology in press Doi:10.1111/mec.15415
この記事を書いた人

-
国立遺伝学研究所 生態遺伝学研究室(北野研究室)所属(日本学術振興会特別研究員PD)
京都大学大学院博士課程修了。博士(理学)。国立遺伝学研究所博士研究員を経て2020年より現職。主にヨシノボリ類やトゲウオ類などの魚類を対象に、進化生態学や進化遺伝学に関する研究に取り組んでいます。特に、DNA配列をもとに進化の歴史を復元することに力を入れています。



