細胞は力刺激を受けると細胞間の接着を強くする? – 力のシグナル伝達による発生制御
発生制御における力
受精卵が卵割を繰り返し、さまざまな性質をもった細胞が生まれ(分化)、個体へと成長する過程「発生」。とくにその初期には、細胞集団は将来の臓器形成に向けて、胚の中をダイナミックに移動して位置を変えます。ある細胞集団が移動すればその運動は別の細胞集団に影響を与えます。細胞は別の細胞に化学シグナルを送ることはよく知られていますが、細胞運動は押したり(圧縮力)、引っ張ったり(引張力)といった「力」という物理シグナルを周囲の細胞に与えるのです。
発生の初期ばかりでなく、成体においても、血管を流れる血液が生み出す力、心臓の拍動が生み出す力など、力は私たちの体のさまざまな場所で生まれています。この半世紀のあいだ、生命科学研究では主に遺伝子やタンパク質の役割に焦点を当てて研究が行われてきました。発生生物学においても生物の形がどのようにできるかなど、謎に富んでいた多くの発生現象の仕組みを「分子の言葉」で解き明かすことに成功してきました。しかし、最近、物理的な要素として力の意義についても注目されるようになったのです。
力に対する細胞の応答
細胞が力の刺激を受けるとどのように変化するのか? この研究は長いあいだ、主に培養細胞を用いて、さまざまな遺伝子の発現変化を解析することによって行われてきました。有名な研究は2006年のDischerらによって報告された、基質の硬さ(弾性)によって間葉系幹細胞の分化方向が決まるという発見です。
私たちは、培養細胞ではなく、初期胚がその形を作り上げる過程(形態形成)において、細胞集団が胚の中で生まれる力にどのように応答するか興味をもち、この一連の研究を行ってきました。まず私たちは遺伝子発現ではなく、タンパク質のリン酸化に着目しました。リン酸化反応は遺伝子発現変化より早く起こるため、細胞外からの刺激に対する迅速な細胞応答を解析できると考えたのです。
また、その細胞応答の全体像を描き出すために、「リン酸化プロテオーム」という方法を用いました。このために、米国プリンストン大学のCristea教授と共同研究を行い、質量分析計という好感度にタンパク質を検出する機器を用いて、検出しうるすべてのタンパク質、ペプチドのリン酸化を捉えることを試みたのです。
また、研究材料は卵の直径が1.2mmほどで比較的大きく、タンパク質の定量的解析のために多くの受精卵が集められるアフリカツメガエルを、初期胚に力を負荷する方法として、多数の受精卵に均一に負荷できる遠心分離機を用いることにしました。
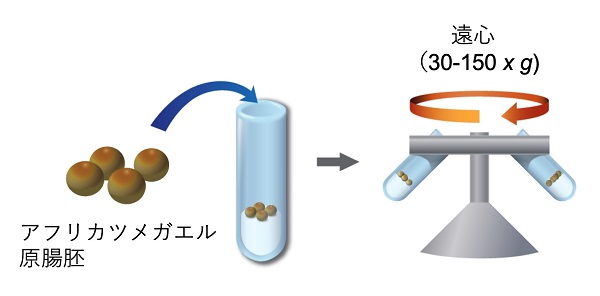
原腸胚をアガロースの上に並べ、遠心によって力を負荷した。短時間(10分程度)の遠心で胚は変形するが、遠心停止後元の球形に戻り正常に発生する。この胚を試料としてリン酸化プロテオームを行った(作図は山梨大学森田仁博士)。
力によって強くなる組織
力によるタンパク質のリン酸化状態の変化に関する膨大なデータを解析したところ、細胞-細胞間接着や細胞-基質間接着に関わると考えられている鍵タンパク質(ZO-1やインテグリン)に機能的に関連するタンパク質群が、力刺激によって迅速にリン酸化され、それらは複雑なネットワークを形成していることがわかりました。
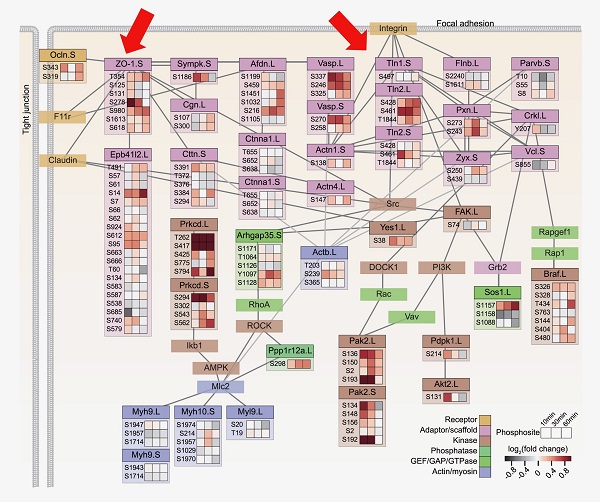
ZO-1(左矢印)およびインテグリンの制御タンパク質Talin (Tln)(右矢印)に機能的に関連するタンパク質群のネットワーク構造が明らかになった。マス目は左から遠心停止後10分、30分、60分の各タンパク質のリン酸化状態を示し、色が濃いほどリン酸化が亢進したことを示す。(Hashimoto et al., Cell Syst., 2019を改変)
また、タイトジャンクション(密着結合)という細胞と細胞のあいだをシールし、体液や分子の移動を防ぐ構造をつくるために必須のタンパク質であるZO-1というタンパク質が、リン酸化に伴って細胞質から細胞膜に移動し、ジャンクション構造に集積することがわかりました。これらの結果から、力刺激を受けた初期胚細胞は細胞-細胞間接着を強化すると考えられます。
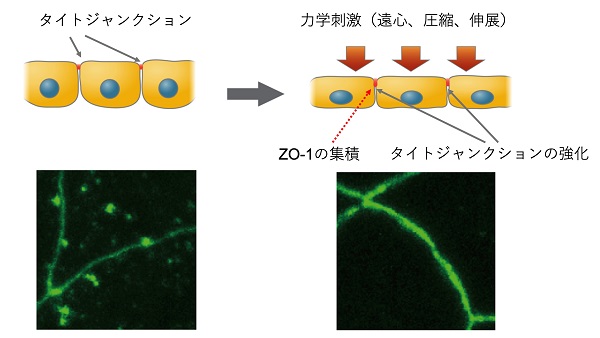
力により細胞は変形し、その結果、ZO-1は細胞質からタイトジャンクションに集積するようになり、細胞-細胞間接着が強化される(上段:細胞を水平に見た模式図)。GFPで標識したZO-1は細胞質に凝集体として存在するが、力刺激によって細胞膜に集まる(下段:細胞を上から見た蛍光顕微鏡像)。
この現象は、上皮細胞が性質を変え運動能を獲得するEMTと呼ばれる現象と逆の現象で、METと呼ばれています。また、F-アクチンが細胞表層に集まり、組織の弾性も高くなることがわかりました。こういった応答は、組織が力によってバラバラになるのを防ぎ、統合性を保って発生を正常に進めるためのフィードバック機構であり、実に理にかなった巧妙な生物のしくみであると解釈しています。また、EMTはガンの浸潤・転移の主要な原因と考えられており、EMT-MET のスイッチング機構の解明はがん研究にも貢献することが期待できます。
力のシグナル伝達経路はあるか?
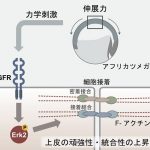
では、細胞はどのように力を感知しているのでしょうか? 私たちの最近の研究で、FGFと呼ばれる細胞増殖因子の受容体が関わっていることが明らかになりました。通常FGF受容体はそのリガンドであるFGFが結合することによって活性化されるリン酸化酵素です。しかし、興味深いことに、この力への応答機構にはFGFは必要ないこともわかりました。
力による細胞-細胞間接着の強化は遠心ばかりでなく、圧縮、伸張によっても起こることから、おそらく力によって細胞が変形することが、FGF受容体を活性化するのではないかと考えています。実際に、原腸形成という胚組織が大きく動き再編成される運動では、胚の表面を覆う外胚葉細胞が大きく伸展している、つまり力を受けていることがわかります。
FGF受容体が活性化した結果、Erkという細胞内因子がリン酸化され活性化することによって、細胞質から核内に移行します。その後、どのような経路をたどってZO-1がジャンクションに集積するのかはまだわかっておらず、それを解明することが私たちの現在の課題です。いずれにしても、力のシグナル伝達経路は既存のシグナル伝達経路を利用して、細胞の変化を引き起こしているようです。
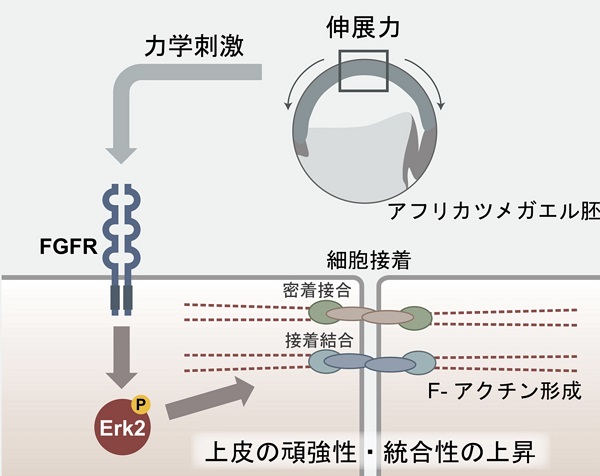
力はFGF受容体、Erk2を活性化し、その後ZO-1はタイトジャンクションに集積する。細胞接着分子カドヘリンも同様の挙動を示し、アドヘレンスジャンクションに集積する。アクチンも表層に集まる結果、組織の統合性、頑強性が高まる (図は基礎生物学研究所プレスリリースより転載)。
私たちはこのシグナル伝達系がマウスにも存在するということを証明するための研究も行っています。もし存在すれば、初期胚細胞の力への応答機構は哺乳動物にまで保存され、生物進化にとっても非常に重要な機構であるといえるでしょう。
力を測る、操る
遺伝子やタンパク質の定量技術は進歩していますが、力は目に見ることができず、とくに生きた生物のなかで力を定量することは至難の技であると考えられています。
力によって細胞が変形し、その結果起こる分子構造変化を、蛍光の変化から捉えるプローブや細胞内に微小なビーズを入れてその流動性から力を推定する方法など、さまざまな測定技術に加えて、ガラスニードルや磁気ビーズを使って細胞に局所的な力刺激を与えるなどの操作技術も開発されつつあります。
今後多細胞からなる生物の複雑な発生過程における力の意義の全貌解明には、非侵襲で力を測定する技術や細胞・組織を操作して摂動を与える技術の開発が鍵になるものと思われます。
参考文献
- Engler AJ, Sen S, Sweeney HL, Discher DE. Matrix elasticity directs stem cell lineage specification. Cell 126, 677-689. 2006
- Hashimoto Y, Kinoshita N, Greco TM, Federspiel JD, Jean Beltran PM., Ueno N*, Cristea IM*. Mechanical Force Induces Phosphorylation-Mediated Signaling that Underlies Tissue Response and Robustness in Xenopus Embryos. Cell Syst. 8, 226-241, 2019 *共責任著者
- Kinoshita N, Hashimoto Y, Yasue N, Suzuki M, Cristea IM, Ueno N. Mechanical Stress Regulates Epithelial Tissue Integrity and Stiffness through the FGFR/Erk2 Signaling Pathway during Embryogenesis. Cell Rep. 30, 875-3888.e3., 2020
この記事を書いた人
-
自然科学研究機構基礎生物学研究所 教授。
1984年筑波大学農学研究科修了(農学博士)。米国ソーク研究所でのホルモン分泌調節因子や細胞増殖因子の研究を経て、帰国後発生生物学における形態形成、とくにBMPによる背腹軸形成の制御機構の研究を行う。この10年ほどは原腸形成や神経管形成など胚組織のメカニクス、力による発生制御機構の研究を行っています。