「微生物ダークマター」全貌解明への道 – これまで誰も分離培養できなかったアーキアを培養してわかったこと
未培養微生物「微生物ダークマター」
私たちの住む地球の表層環境は、微生物で満ちています。微生物のいないところを探すほうが難しいといってもいいでしょう。それでは、そんな微生物のことを私たちはどれだけよく知っているのでしょうか。実は、その全貌はほとんど明らかになっていないのです。
地球上に存在する全微生物の種の数は数百万種以上と推定されておりますが、その99%以上がまだ実験室内で培養できていません。培養できないと、その微生物の生き様(増殖できる条件や速度、保有する機能、生産性、生態系の中での役割など)を明らかにすることはできませんし、その微生物を有効利用することもできません。たった1%以下のすでに培養できている微生物だけみても多様な機能を有しており、食・医療・環境保全といったさまざまな場面で私たちの生活を支えてくれています。
もっと面白くてもっと有用な微生物が自然界には存在するに違いないのですが、それらは未発見・未利用のまま残されているのです。そういった未だ培養できてきない微生物は「微生物ダークマター」とも呼ばれ、多くの研究者から注目を集めています。
温泉に生息する未培養アーキア系統群「THSCG」
環境中のDNAを解読することで、培養しなくてもどんな系統の微生物がそこに存在するのか調べることができます。そのような培養に依存しない分子生物学的手法によって、「微生物ダークマター」はさまざまな自然環境に存在することが明らかにされてきました。日本人に馴染みの深い温泉も、その生息域のひとつです。
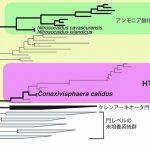
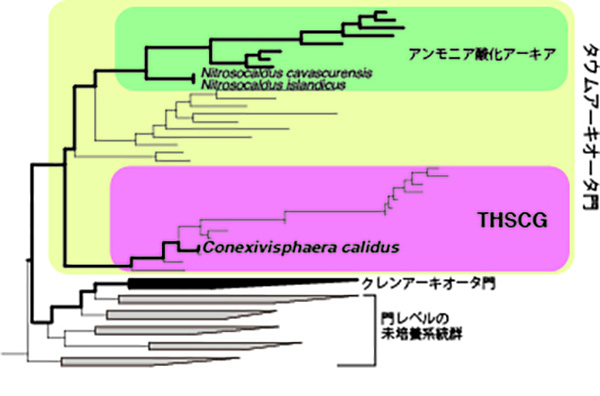
特に温泉では、真核生物でもバクテリアでもない、第三の生物「アーキア(古細菌)」が優占して存在することが多々あります。温泉アーキアも未培養の系統を多数含みますが、そのひとつに、1999年に日本人の研究者によって初めて報告されたTHSCG(Terrestrial Hot Spring Crenarchaeotic Groupの略)という系統群があります。
THSCGは、当時はクレンアーキオータ門に分類されておりましたが、現在では、2008年に新たに提唱されたタウムアーキオータ門に分類されており、同門のなかで最も古くに分岐した系統です。タウムアーキオータ門のなかで培養できている種はすべて、アンモニアを酸化してエネルギーを得て生育する種でした。タウムアーキオータ門はアーキアのなかで唯一、アンモニアを酸化する種を含みます。
一方で、これまでの環境DNA解析から、THSCGアーキアはアンモニア酸化ではなく、有機物を分解して硫黄や硝酸を還元することでエネルギーを得て生育する種であると予想されました。しかしながら、THSCGアーキアの分離培養に成功した例はなく、その生理機能や生態学的な役割を裏付ける証拠は得られていませんでした。
THSCGアーキアの分離培養に至るまで
私たちの研究チームは、世界で初めてTHSCGに属するアーキアの分離培養に成功しました。
以前の研究によって、栃木県奥塩原温泉にTHSCGアーキアが生息することがわかっていましたので、まず同温泉の灰色に濁った高温酸性の温泉水(57ºC, pH 2.2)を採取しました。採取した温泉水を、エネルギー源となる物質(硫黄や鉄、水素、有機物など)を含む培地に添加して、さまざまな温度・pH条件下で培養しました。約1か月間培養した後、70ºC、pH 5.0で培養したFe(III)と有機物を含む培養液を注意深く顕微鏡観察したところ、一般的な微生物細胞サイズ(1-2 µm程度)よりも小さな微生物が増殖していることが確認できました。この培養液のDNA解析の結果、THSCGアーキアが含まれることがわかりました。
この培養液をもとにして、限界希釈法(微生物細胞が1個含まれるかどうかいうところまで希釈して培養する方法)を何度か繰り返すことで、THSCGに属するアーキアを純粋分離することに成功しました。この分離株をNAS-02株と呼ぶことにし、さらに新属新種であることが明らかだったため、暫定的にConexivisphaera calidus(以下、C. calidus)と名付けました。この名前は、「熱いところに住む(calidus)、クレンアーキオータ門とアンモニア酸化アーキア群との間をつなぐ(Conexivi)球菌(sphaera)」に由来します。
C. calidusは、60ºCから70ºCの間で生育する好熱菌で、pH 4.5から5.5の弱酸性条件下で生育します。この非常に狭い範囲内の温度・pH条件下でしか生育できないという生育特性が、これまでC. calidusが培養されてこなかった理由のひとつとして挙げられます。
C. calidusの細胞は、直径約0.5 μmのとても小さい不定形球菌です。400倍程度の倍率で顕微鏡を覗いてみても、注意深く観察しないとゴミと見間違うレベルです。増殖速度は非常に遅く、一番よい条件で増殖するときも1回分裂するのに約40時間かかります(ちなみに、実験でよく使われる大腸菌は約20分で分裂します)。さらに、1か月間培養しても最大で107 cells/ml程度の菌密度にしかならないので、培養液の濁りも目視ではまったくわかりません。この小さい細胞サイズと低い菌密度も、これまでC. calidusが培養されてこなかった理由のひとつでしょう。
そんな扱いづらい本菌株の分離を実際に担当したのは、当時東洋大学の大学院生であった大西真史くんです。また、その後の生理性状解析は、同大学の大学院生であった長森麻衣さんによって行われました。この2人の大学院生の粘りと努力によって、これまで謎に包まれていたTHSCGアーキアの生理・生態・進化への道が開けたのです。

THSCGアーキアを培養できた決め手
これまで誰も分離培養できなかったC. calidusを、本研究で培養できた決め手は何でしょうか。
そのひとつは、pH 5.0で培養を試みたことです。元の温泉水のpHは2.2でしたが、本研究では、たまたまpH 5.0の条件でも培養を行ったため、結果的にpH 4.5から5.5で増殖できるC. calidusが培養できたのです。温泉水に含まれる泥の粒子の中には、pHが5付近になっている微小空間があり、そこにC. calidusは生息していたと予想しています。培養条件をひとつ増やすということは、実験の手間が増えることになりますが、その手間を惜しまずに培養を試みたことで、思いがけずC. calidusの分離培養に繋がったのです。
もうひとつの決め手は、Fe(III)を含む培地を用いたことです。C. calidusは、エネルギー源および炭素源としてタンパク質を含む基質(酵母エキスやペプトン)を利用し、電子受容体としてFe(III)を還元して生育します。1998年にNature誌に掲載された論文で、Fe(III)を還元するとは夢にも思われていなかった好熱菌の多くが、実はFe(III)還元菌であったと報告され、論文中には「未培養微生物の分離には、Fe(III)を含む培地を用いた培養法が有効かもしれない」と記されていました。この予言は正しかったのです。
ただし、現時点では、それらの好熱性Fe(III)還元菌(C. calidusを含む)が、どうやってFe(III)を還元しているのか、そのメカニズムは未だ明らかになっておらず、鍵となる遺伝子も見つかっていません。つまり、いくら環境DNA解析で培養を介さずにTHSCGアーキアのゲノムを再構築したとしても、そのゲノムに由来する微生物がFe(III)還元菌かどうかはわからないのです。ここに培養に依存しない環境DNA解析の限界があります。コッホの時代から100年以上受け継がれてきた「分離培養」は、今もなお微生物学において非常に有効な手法なのです。
THSCGアーキアが培養できてわかった多くのこと
分離培養できたことでわかったことが、他にもいろいろあります。たとえば、C. calidusは、Fe(III)の代わりにチオ硫酸や元素硫黄も還元できることがわかりました。全ゲノム解析の結果、C. calidusのゲノムには、硫黄還元に関わる遺伝子がいくつか含まれていることも明らかになりました。興味深いことに、それらの遺伝子は、温泉に生息するクレンアーキオータ門の好熱菌から水平伝搬で獲得されたものであることが、系統解析によって示されました。温泉水には鉄の他にも硫黄が豊富にふくまれますので、C. calidusは硫黄も利用できるように進化して、他の微生物との生存競争に負けないようになったのではないかと推定されます。
また、C. calidusはタンパク質を含む基質を分解して生育することから、温泉環境では他の生物の死骸を分解する「スカベンジャー」としての役割を果たしていると考えられます。
さらに、C. calidusはタウムアーキオータ門のなかでアンモニアを酸化「しない」種の初めての報告例です。一方で、タウムアーキオータ門のアンモニアを酸化「する」種のなかで最も古い系統であるNitrosocaldus属も、温泉に生息する好熱菌です。これらのことは、「アンモニア酸化アーキアは温泉で誕生した」という説を強く支持しています。
まとめ
本研究では、「微生物ダークマター」の一部であったTHSCGに属するアーキア、C. calidusを世界で初めて分離培養することに成功しました。これまで謎に包まれていたC. calidusの生理性状が明らかになり、その進化の過程や生態系のなかでの役割にも言及できるようになりました。
C. calidusの分離培養に至るまでは、先行研究の精査はもちろん、根気よく培養を続ける「粘り」、そして「運」が必要でした。重要なことは、これまでC. calidusを培養できなかった理由が、今回培養できたことで初めて見えてきた、ということです。分離培養を地道に続けて、成功例を積み重ねることで、また新たな「微生物ダークマター」の培養への道が拓けると期待されます。
そうしていつの日か「微生物ダークマター」の全貌が明らかになったとき、きっと世界は変わります。食糧難・病気・環境汚染・エネルギー枯渇など人類が直面する多くの問題を、微生物が解決してくれることでしょう。また、その機能を獲得するまでの進化の過程を明らかにすることで、人類が求める新機能・高機能を自在にデザインできるようになるかもしれません。
分離培養は、古典的・地味・煩雑・リスキー等々ネガティブな印象を持たれることが多く、実際その通りなのですが、それでも世界を変える大きな可能性を秘めています。今後もコツコツと分離培養を進めて、「微生物ダークマター」の全貌に迫っていきたいと考えています。
参考文献
- Kato S, Itoh T, Yuki M, Nagamori M, Ohnishi M, Uematsu K, Suzuki K, Takashina T, Ohkuma M. 2019. Isolation and characterization of a thermophilic sulfur- and iron-reducing thaumarchaeote from a terrestrial acidic hot spring. ISME J 13:2465-2474.
- Takai K, Horikoshi K. 1999. Genetic diversity of archaea in deep-sea hydrothermal vent environments. Genetics 152:1285-1297.
- Vargas M, Kashefi K, Blunt-Harris EL, Lovley DR. 1998. Microbiological evidence for Fe(III) reduction on early earth. Nature 395:65-67.
この記事を書いた人

-
理化学研究所・バイオリソース研究センター・微生物材料開発室
2009年東京薬科大学大学院生命科学研究科博士課程後期修了。博士(生命科学)。同大学嘱託研究員、理化学研究所基礎科学特別研究員、デラウェア大学客員研究員(日本学術振興会海外特別研究員)、海洋研究開発機構特任研究員などを経て、2018年4月から現職。好熱性アーキアや鉄酸化・還元細菌などの難培養微生物の生理・生態・進化についての研究を進め、さらにそれらの微生物のリソース化を目指している。