X精子だけがもつタンパク質TLR7/8の発見と、雌雄の簡単な産み分け方法
ほ乳類の性別が決まる仕組み
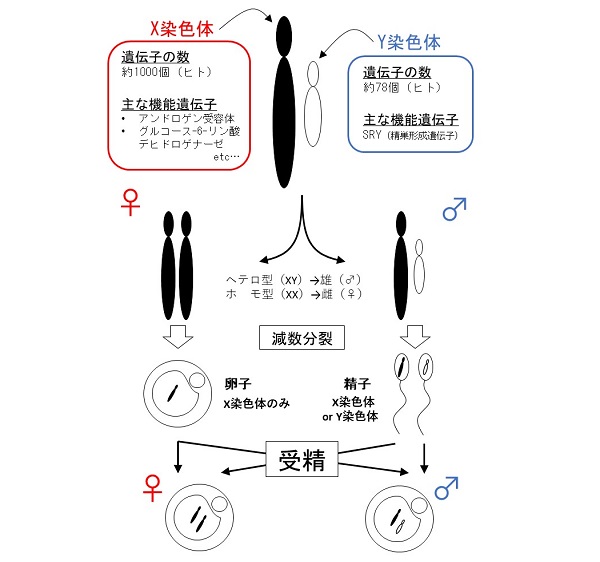
ほ乳類において、生まれてくる子どもの性は、性染色体と呼ばれるX染色体とY染色体の組み合わせで決まります。具体的には、X染色体を2つもつと雌(XX)になり、X染色体とY染色体をもつと雄(XY)になります。
精巣や卵巣では、減数分裂と呼ばれる方法で子どもを作る源となる配偶子(厳密にいうとほ乳類では、半数体となる雌性配偶子(卵子)が形成されるときには精子が侵入しているため、雌性配偶子は存在しない)が作られることから、卵子はX染色体を、雄の配偶子である精子はX染色体あるいはY染色体のいずれかをもちます。
このように作られた両配偶子(卵子と精子)が出会い、生命のスタートである“受精”が起こることから、子どもが授かる性は、受精する精子がもつ性染色体がX染色体であれば雌(XX)となり、Y染色体であれば雄(XY)となります。したがって、精子がもつ性染色体を見分けることができれば、生まれてくる子どもの性別を選択できるといえます。
精子の能力を均一化する精子細胞間の「ブリッジ構造」
染色体には、細胞内でさまざまな機能を発揮し、細胞が生存するために必須なタンパク質をコードする遺伝情報(遺伝子と呼ぶ)が記録されています。しかしながら、X染色体には、男性ホルモンを認識するアンドロジェン受容体や、細胞の生存に必須なグルコース-6-リン酸デヒドロゲナーゼなど、1000個以上の遺伝子が記録されている一方で、Y染色体には精巣を形成するのに必須なSRY以外に機能的な遺伝子はほとんどなく、遺伝子の数は80個程度しかありません。
つまり、X染色体にコードされた遺伝子がX染色体をもつ精子(X精子)においてのみ発現していれば、X精子とY染色体をもつ精子(Y精子)に機能差が生まれ、それが発揮される環境では、雌雄比率が異なることになります。しかしながら、雌雄比率は多くのほ乳類でほぼ1:1であることから、X精子とY精子のあいだに機能の違いはないと考えられてきました。
異なった染色体構成にもかかわらず両者に機能差が発揮されないカギは、精子が作られる過程に存在する「ブリッジ構造」にあります。ブリッジ構造は、細胞間架橋とも呼ばれる構造で、その名のとおり繋がった細胞の間でタンパク質や遺伝子を橋渡し(共有)する役割をもちます。
精子は、精子幹細胞が減数分裂を経て精子細胞へと分化した後、大きく形態を変えて(変態して)産生されます。この精子完成前の精子細胞(X染色体あるいはY染色体をもつ2種類が存在する)の段階では、ブリッジ構造によりX精子細胞とY精子細胞が繋がれています。つまり、XあるいはY染色体のみをもつように変化した精子細胞間でタンパク質や遺伝子が共有されるので、ブリッジ構造を介してX染色体をもたないY精子であっても、X染色体由来のタンパク質や遺伝子が得ることができ、Y精子でもX精子と同様の機能をもつことが可能になります。
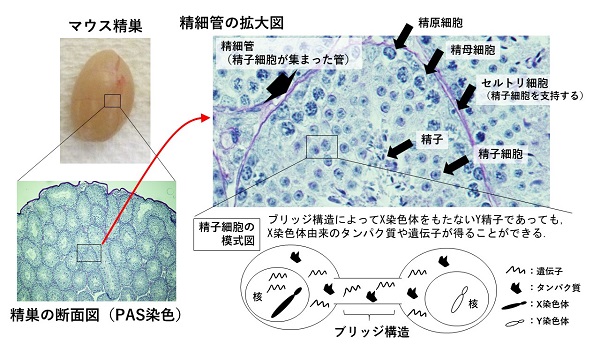
さらに、ブリッジ消失後の精子細胞と精子では、染色体の構造が大きく変化するので、遺伝子発現は生じないことも報告されています。このようなシステムによって、X精子とY精子の機能的な選別が起こらず、大部分のほ乳類において生まれてくる子どもの性比(雄と雌の割合)は1:1になると考えられてきました。
X精子にしか発現しない「Toll様受容体7/8(TLR7/8)」の発見
このような通説が存在する一方で、我々は-196℃という環境で凍結したブタ精子を人工授精に用いた場合、凍結していないブタ精子を用いた場合と比較して雌雄の割合が大きく偏るという興味深い知見を得ました。ブタは、ヒトやウシなどと異なり、一度のお産で10匹以上の産子が誕生しますので、そのお産ごとの雄雌比率を算出した結果です。
このことは、-196℃という通常さらされない環境を経た精子では、X精子とY精子に機能差が発揮された可能性を示しています。そこで、機能差は存在するという異端な視点で、マウスの精巣を詳細に解析した結果、精子細胞間のブリッジ構造が精子細胞から精子に変態する過程で消失した直後は、精子細胞は遺伝子発現を行っていることを突き止めました。
これらの結果から、ブリッジ構造が消失した後にX染色体から発現する遺伝子により、X精子とY精子のあいだに潜在的な機能の違いが作られ、それは通常の受精環境では覆い隠されていますが、特殊な環境下で発揮されるのではないかと考えました。
この仮説を立証するため、まずマウス精子に含まれる全遺伝子の解読を行い、(1) X染色体にコードされているもの、(2) 環境を認識する受容体をコードするもの、そして(3) ブリッジが切断した後に発現するもの、という3段階のスクリーニングを行いました。
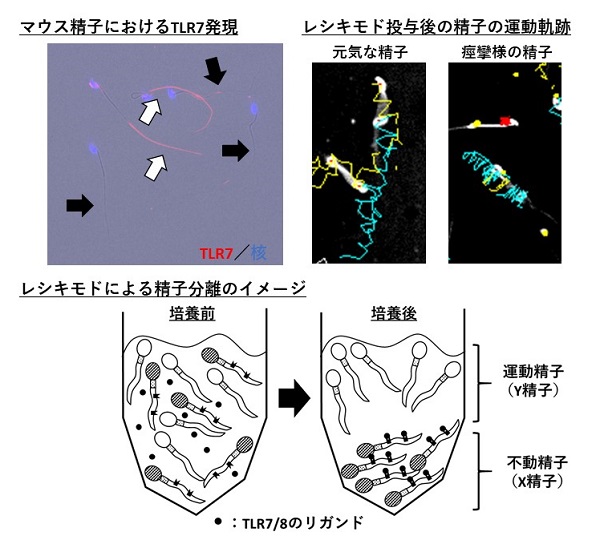
すると、Toll様受容体という病原体を感知して排除する自然免疫を作動させる受容体の7番(TLR7)と8番(TLR8)が候補化されました。そこで、マウス精子においてTLR7/8の分布を見てみると、それはX染色体をもつ精子にしか発現していないことを確認しました。このような戦略によって、我々は世界で初めてX精子とY精子で異なる発現を示すタンパク質を見つけ出しました。
TLR7/8活性化が簡単な雌雄産み分けを可能にする
次に、X精子にしか存在しないTLR7/8がX精子とY精子のあいだで機能差を生み出すのかを明らかにするため、TLR7/8を活性化させる薬剤であるレキシモドを加え、マウス精子の挙動を観察しました。すると興味深いことに、半数の精子は元気に動いている一方で、半数の精子はビクビクと痙攣をしたかのような動きを示しました。
元気に動く精子には、重力に逆らって上に向かって泳ぐ性質があることから、レキシモドを加え泳がせた精子の上層と下層を分け、それぞれ体外受精試験に供試しました。すると、驚くべきことに上層精子を用いると9割以上がXY胚(雄)となり、下層精子では8割程度がXX胚(雌)となることが明らかとなりました。つまり、X精子しかもたないTLR7/8を特殊な環境で活性化させると、X精子とY精子の分離、さらには雌雄の産み分けが可能になることが明らかとなりました。
では、本研究に用いたマウス以外ではどうでしょうか? Web上に公開されているゲノムのデータベースを見てみると、マッコウクジラ、オポッサム、カモノハシを除くすべてのほ乳類において、TLR7はX染色体上にコードされていました。すなわち本研究で明らかにした技術は、ほ乳類全体に適用でき得る可能性を有しています。この可能性はヒトにおいても同様なことがいえますが、倫理的観点より研究対象に含めていません。
現在では、産業家畜、特にウシやブタでの実用化を目指して、研究を進展させています。雌雄の産み分けについて、ウシではセルソーターという高額な特殊機器を用いてX精子とY精子を分離することに成功していますが、問題点として、その分離過程において精子の受精する能力が低下してしまうことや、多量の精子が無駄になってしまうことがあり、より高効率な手法が求められています。またブタでは、セルソーターによる分離は適用できておらず雌雄産み分け自体ができていません。
そこで、第一にウシやブタの精子でもTLR7が発現しているか否か検討したところ、マウスと同様にTLR7は半数の精子にしか存在していませんでした。さらに、レキシモド処理を用いることで、ウシ体外受精では90%以上の確率で雄胚を作出することに成功し、ブタ人工授精では70%程度の確率で雄個体を産ませることに成功しました。
本研究がもつ可能性
興味深いことに、B型肝炎ウィルスに感染した患者は男児を産みやすいという疫学的な報告があります。 B型肝炎ウィルスは、TLR7/8と強い関係性をもつことが報告されており、TLR7/8の存在が病態による雌雄の偏りの原因である可能性が示唆されます。またアカシカでは、母親の栄養状態によって子どもの雌雄が決定されることが知られており、TLR7/8以外にも存在するX精子の隠れた機能によって差異がもたらされているかもしれません。
本研究で行っているX精子とY精子の違いに関する研究は、まだ始まったばかりであり、明らかとなっていないことは数多く残っています。今後は、本研究をスタートとして、マウスで確立した雌雄産み分け法をより効率化し、その他のほ乳類に実用化していくだけでなく、なぜX染色体にだけ発現するタンパク質があるのか? なぜX精子とY精子に違いが存在するのか? など、進化的側面を含めて明らかとなっていない謎の解明に取り組んでいきたいと考えています。
参考文献
Umehara, T., Tsujita, N., & Shimada, M. “Activation of Toll-like receptor 7/8 encoded by the X chromosome alters sperm motility and provides a novel simple technology for sexing sperm” PLoS biology, 17(8) (2019) DOI: 10.1371/journal.pbio.3000398
この記事を書いた人
-
梅原 崇
広島大学大学院統合生命科学研究科 家畜生産機能学(家畜生殖学)研究室 助教。
愛媛県出身。広島大学大学院生物圏科学研究科博士課程修了。博士(農学)。雌雄両性の生殖細胞(精子や卵)と、その成熟を制御する生殖器(精巣や卵巣)に興味をもち、これら機能やメカニズムの解明・理解を通して、畜産業の発展に繋がる技術を開発することを目指しています。特に、現在では本記事で紹介した“雄の生殖細胞である精子がもつ秘められた能力に着眼した新たな雌雄産み分け法”とともに、マウスをモデルとした“病態や加齢に伴って低下する雌の繁殖能力の原因探索と、その予防法や治療法の開発”といった2つのテーマで研究しています。
辻田 菜摘
広島大学大学院統合生命科学研究科 家畜生産機能学(家畜生殖学)研究室 研究員。
長崎県出身。広島大学大学院生物圏科学研究科博士課程後期修了。博士(農学)。動物の生命現象を生体内の遺伝子やタンパク質などの発現量の変化から解き明かすことの面白さに興味を持ち、分子生物学系の研究室で魚類の性転換やクラゲの変態について研究をおこなってきました。2018年秋より現在の研究室で研究員として働き始め、雌雄産み分け法の改良につながる研究を進めています。
島田 昌之
広島大学大学院統合生命科学研究科 教授。
1999年 広島大学助手、2003年 博士(獣医学)取得、2004~2005年 米国ベイラー医科大学 客員研究員、2006年~2016年 広島大学 准教授、2017年~ 広島大学 教授。専門分野は家畜繁殖学、生殖生物学。マウスをモデルとして、雌雄の生殖機構を分子生物学的、内分泌学的に解析をする基礎研究から、日本の豚肉生産の約半数を支えている人工授精技術の開発などの実用化研究までを行っている。
この投稿者の最近の記事
 研究成果2019年12月24日X精子だけがもつタンパク質TLR7/8の発見と、雌雄の簡単な産み分け方法
研究成果2019年12月24日X精子だけがもつタンパク質TLR7/8の発見と、雌雄の簡単な産み分け方法


