多様な食物に適応する仕組みとは? – ショウジョウバエ近縁種間の差を生む”炭水化物応答機構”
広食性種が持つ適応システムとは?
栄養は、生物の成長や生命の維持にとって不可欠です。進化の過程で、動物は周囲の栄養環境と相互作用しながら、種ごとに異なる多様な食性を獲得してきました。この食性の幅の違いから、大きく分けて2種類の動物が存在します。さまざまな物を食べることのできる広食性種(generalist)と、特定の物だけを食べて生きる狭食性種(specialist)です。
これまで、狭食性種が特定の食物を好んだり、食物に含まれる毒に耐性を持ったりする仕組みについては、研究が比較的進められてきました。一方で、ヒトを含めた広食性種がどのようにしてさまざまな栄養環境に柔軟に応答し、適応を実現しているかについては不明な点が多く残されています。
分子生物学において普遍的な生命現象の研究に用いられてきたモデル生物であるキイロショウジョウバエは、自然界では全世界の人家近くに生息し、さまざまな発酵した果物や野菜を食べる広食性種です。このモデル生物には、森や砂漠、渓流などの環境に特化した種を含む 2,000以上の近縁種が存在し、なかには、限られた地域で特定の発酵した植物や果実だけを食べる狭食性種もいます。
たとえば、セイシェルショウジョウバエはインド洋のセイシェル諸島にのみ生息し、ノニ(ヤエヤマアオキ)の果実だけを餌とする狭食性種であることが知られています。そこで私たちは、このように自然界での食性が異なるショウジョウバエの近縁種間で、栄養に対する適応能力や生体応答の比較解析を行うことにより、広食性種が多様な栄養環境に適応できるメカニズムの解明を目指しました。
栄養への適応能力をショウジョウバエ近縁種間で比較
まず、私たちは、キイロショウジョウバエを含む広食性2種と、自然界では特定の果実や花のみを食べる狭食性3種を用いて、異なる栄養環境に対する幼虫期の適応能力が種間でどのように異なるかを調べました。これらの種は、ゲノム情報が整備されているため、栄養への応答を比較解析するうえで活用できるのではないかと考えたからです。
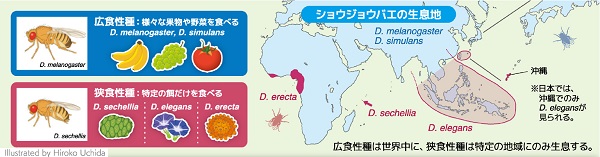
D. melanogaster: キイロショウジョウバエ、D. simulans: オナジショウジョウバエ、D. sechellia: セイシェルショウジョウバエ、D. elegans: カザリショウジョウバエ、D. erecta: エレクタショウジョウバエ
栄養環境の違いを捉えるうえで着目したのが、タンパク質(Protein)と炭水化物(Carbohydrate)の比率(P:C ratio)で栄養バランスを変化させる方法です。これまでに、食餌中のP:C ratioの違いが、昆虫から哺乳類の寿命や生殖能力などに大きな影響を与えることが示されています。たとえば、キイロショウジョウバエやマウスの寿命は、どちらも炭水化物の比率が高い食餌条件下で長くなることが報告されています。
そこで、同一カロリーで栄養バランスの異なる3種類の実験餌(炭水化物の比率が高い餌、炭水化物とタンパク質の比率が中間程度の餌、タンパク質の比率が高い餌)を作製し、各種の幼虫を孵化直後からこれらの餌で飼育して、その成長を比較しました。
その結果、広食性種の幼虫はどの実験餌においても正常に成長できるのに対し、狭食性種はいずれも炭水化物の比率が高い餌では蛹にまで発生できないことを見出しました。特に、狭食性種のセイシェルショウジョウバエとカザリショウジョウバエは、餌中の炭水化物の比率が高くなるにつれて、蛹までの発生率が顕著に低下する傾向を示しました。
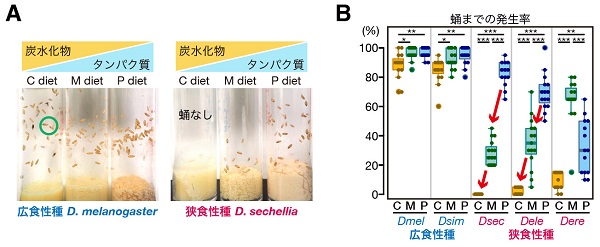
(A) 栄養バランスが異なる3種類の実験餌で育てた、広食性種のキイロショウジョウバエと狭食性種のセイシェルショウジョウバエ。広食性種キイロショウジョウバエは、どの餌条件でも蛹(緑色の丸)にまで発生できるが(左)、狭食性種セイシェルショウジョウバエは炭水化物の比率が高い餌(図中 “C diet”)で成長できない(右)。
(B) ショウジョウバエの近縁5種について各実験餌における蛹までの発生率を定量化した結果。狭食性種のセイシェルショウジョウバエ(図中 “Dsec”)とカザリショウジョウバエ(図中 “Dele”)は、餌の炭水化物比率が高くなるにつれて、発生率が低下する(赤矢印)。狭食性種のエレクタショウジョウバエ(図中 “Dere”)の発生率は、広食性種及び上記の狭食性2種とは異なる。
そこで、狭食性種の発生率の低下にとって鍵となる食餌中の成分を調べるために、各種の食性の違いに着目し、これらの近縁5種の幼虫が自然界で食べている餌の栄養成分を比較しました。日本国内に生息する広食性種と狭食性種の餌については、実際に野生の幼虫が食べていた発酵した果物や花を採取し、ガスクロマトグラフィー質量分析法によりその成分を解析しました。

自然界で野生の広食性種キイロショウジョウバエが食べていたバナナ(上段)と、狭食性種カザリショウジョウバエが食べていたアサガオの花(下段)の発酵前と発酵後の写真。アサガオの花については、雄しべや雌しべ、花粉(角括弧)を集めて解析に用いた。矢印は幼虫を表す。
くわえて、文献やデータベースから得られた情報を基にして、各種が野外で食べる植物の栄養バランスの比較を行いました。その結果、広食性のキイロショウジョウバエが野外で食べる餌には高炭水化物食(リンゴやバナナなど)から低炭水化物食(トマトやズッキーニなど)まで幅がある一方、狭食性種セイシェルショウジョウバエの餌であるノニの果実やカザリショウジョウバエの餌であるアサガオなどの花は、どちらも低炭水化物食であることがわかりました。
種間の適応能力の差を生む「炭水化物応答機構」
これらの結果から、自然界で低炭水化物食のみを食べる狭食性種では、食餌中の炭水化物の比率の増加に適切に応答するメカニズムが働いていないのではないかと考えました。そこで、広食性種のキイロショウジョウバエにおいて、摂取した炭水化物に応答して脂肪組織から分泌されるタンパク質TGF-β/Activinと、その下流で制御されるシグナル伝達経路に着目しました。
キイロショウジョウバエにおけるActivinファミリーのひとつであるDawdle遺伝子の変異体は、狭食性種のセイシェルショウジョウバエやカザリショウジョウバエと同様に、実験餌中の炭水化物比率の増加に伴い蛹までの発生率の低下を示しました。
さらに、次世代シーケンサーを用いて遺伝子発現をゲノム規模で解析するRNA-seq解析や、ガスクロマトグラフィー質量分析法を用いて多数の代謝産物量を一度に比較分析するメタボローム解析によって、遺伝子発現および代謝産物の網羅的比較を行いました。その結果、狭食性種セイシェルショウジョウバエでは、TGF-β/Activinシグナル伝達経路を含む、摂取した炭水化物への応答を制御する仕組みである「炭水化物応答制御機構」に異常がある可能性が示唆されました。
広食性種の野生型は、炭水化物応答制御機構を介して、筋肉や腸、脂肪組織など、全身の各組織において200以上ものさまざまな代謝酵素遺伝子の発現量を調節し、異なる餌条件下でも代謝の恒常性を維持できるのに対し、狭食性種セイシェルショウジョウバエと広食性種キイロショウジョウバエのDawdleの変異体ではこのような制御が働かず、高炭水化物条件下で代謝酵素遺伝子群の発現上昇と、多数の代謝産物量の増加を示すことがわかりました。
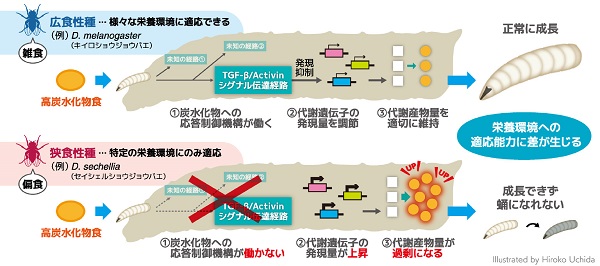
本研究では、遺伝的に近縁ながら自然界での食性が異なるショウジョウバエの種間で、栄養バランスの異なる餌に対する適応能力や、遺伝子発現量および代謝産物量の制御に違いがあることを見出しました。そして、これらの種間の違いは、TGF-β/Activinシグナル伝達経路を含む炭水化物応答制御機構が広食性種では機能しているのに対して、自然界で低炭水化物食のみを食べる狭食性種では、進化の過程でこの機構が失われ機能していないために生じた可能性が強く示唆されました。
おわりに
本研究で用いたショウジョウバエとヒトとのあいだでは、組織やホルモンなどの制御因子の多くが共通しています。加えて、私たちヒトにおいても、肥満や糖尿病などの罹りやすさが人種や個体間で異なることが知られています。今後、本研究での成果や種間比較解析のアプローチを応用することで、個体間、そしてショウジョウバエ以外の生物種間においても、栄養やその他の環境因子に対する適応や生体応答の違いを生み出すメカニズムの研究が広く進むことが期待されます。
また、本研究では、食餌中の栄養バランスの違いが、成長段階にある幼虫期の個体に与える影響に注目し研究を進めてきました。一方で、成長期に摂取した栄養バランスの違いが、成虫にまで発生し加齢していく後期ライフステージにおいても、個体の健康状態に違いを生み出す可能性は十分に考えられます。今後は、本研究で得られた知見を基盤として、成熟した個体の寿命や運動機能、生殖能力などにも着目し、栄養環境の違いが動物の一生に与える影響をさまざまな角度から調べていく予定です。
一方で、現時点では、高炭水化物食に適応できない狭食性種が、炭水化物応答制御機構のどの段階に異常や変異を持つかについて明らかにできていません。その候補には、今回着目した TGF-β/Activinシグナル伝達経路やその上流因子に加え、炭水化物応答制御機構として働く未同定の因子やシグナル伝達経路など、複数の可能性が考えられます。今後、キイロショウジョウバエ以外の近縁種においても、さまざまな遺伝子組み換え系統を利用することが可能となれば、高炭水化物食に適応できない狭食性種の原因究明と、広食性種の適応能力を支える遺伝子ネットワークの解明に近付くと期待しています。
参考文献
Kaori Watanabe, Yasutetsu Kanaoka, Shoko Mizutani, Hironobu Uchiyama, Shunsuke Yajima, Masayoshi Watada, Tadashi Uemura, and Yukako Hattori “Interspecies Comparative Analyses Reveal Distinct Carbohydrate-Responsive Systems among Drosophila Species”(種間比較解析により明らかにされたショウジョウバエ近縁種群における炭水化物応答制御機構の違い)Cell reports 28, 2594–2607 (2019)
DOI:https://doi.org/10.1016/j.celrep.2019.08.030
この記事を書いた人
-
渡辺 佳織
京都大学大学院生命科学研究科特定研究員。博士(生命科学)。2019年京都大学大学院生命科学研究科博士後期課程研究指導認定退学。
上村 匡
京都大学大学院生命科学研究科教授。理学博士。1987年京都大学大学院理学研究科修了。カリフォルニア大学サンフランシスコ校博士研究員、京都大学理学部助手、京都大学大学院生命科学研究科助教授、京都大学ウイルス研究所教授を経て現職。
服部 佑佳子
京都大学大学院生命科学研究科助教。博士(生命科学)。2012年京都大学大学院生命科学研究科博士後期課程研究指導認定退学。京都大学大学院生命科学研究科教務補佐員、特定助教を経て現職。



