植物がもつ再生能力の秘密 – 分化細胞を幹細胞へと変化させる”ステミン遺伝子”の発見
植物がもつ再生能力の秘密
私たち人間をはじめ、動物も植物も、ひとつの細胞である受精卵が分裂をくり返し、増えた細胞がいろいろな性質を持ち特殊化する(分化する)ことで、体ができあがります。ところが、ひとたび分化した細胞でも、さまざまな種類の細胞を作り出すことができる幹細胞へと変化させることができます。
幹細胞についてもう少し説明すると、幹細胞とは、細胞分裂によって自分と同じ幹細胞と、自分自身とは異なった、より特殊化した細胞になる細胞を作り出す細胞のことです。ヒトを含む哺乳類では、通常の状態では分化した細胞が幹細胞になることはありませんが、分化細胞に複数の遺伝子を働かせることで、幹細胞であるiPS細胞に変化させることができます。
ところが植物は、特殊な遺伝子操作をしなくても、挿し木や葉刺しに見られるように、一部の組織から容易に新しい植物体全体を再生させることができます。これは傷害に反応して、茎や葉を作っている一部の分化細胞が幹細胞へと変化し、その幹細胞の働きにより新しい組織や器官ができるようになるためです。
このように植物には、動物と異なり分化細胞を幹細胞へと変化させる能力がもともと備わっています。では、植物はどのような仕組みで分化細胞の性質をリセットさせ、幹細胞へと変化させるのでしょうか?
幹細胞化を誘導する「ステミン遺伝子」の発見
これまでにシロイヌナズナなどの被子植物を用いた研究から、ひとつの遺伝子を葉細胞に働かせることで細胞分裂を再開させ、幹細胞を含む分裂組織を作り出すことが可能でした。しかしながら、細胞分裂を繰り返す一部の分化細胞が幹細胞へと変化するため、幹細胞になる細胞の特定が難しく、分化細胞を幹細胞へと変化させる仕組みについてはよくわかっていませんでした。
そこで私たちは、再生能力の高いコケ植物であるヒメツリガネゴケに着目しました。ヒメツリガネゴケは、胞子が発芽すると原糸体幹細胞が形成され、分裂と細胞伸長を繰り返すことで、糸状の組織である原糸体を作り出します。さらに、原糸体幹細胞から生じた原糸体細胞から茎葉体幹細胞が作り出され、葉をもった構造体である茎葉体が作られます。ところが、茎葉体から葉を切り離して水につけておくと、切断面に面した葉細胞が原糸体幹細胞へと変化します(幹細胞化)。その後、新たにできた原糸体幹細胞が発生を続け、新たな個体を作り出します。
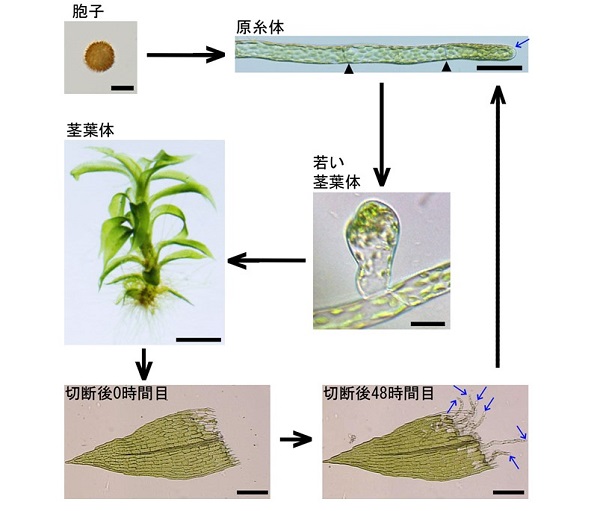
このようにヒメツリガネゴケは、容易に幹細胞化を誘導することができるとともに、幹細胞化する細胞を特定することができます。そのため、分化細胞から幹細胞へと変化する仕組みを調べるのに優れた研究材料です。
私たちの研究チームでは、ヒメツリガネゴケの全遺伝子のなかから幹細胞化に関わる遺伝子を探索し、15個まで絞り込んで個別に機能を調べました。そして、たったひとつの遺伝子を無傷の葉細胞に働かせるだけで、葉細胞を直接幹細胞へと変化させることができることを発見しました。幹細胞へと変化させることができるこの遺伝子を、私たちは「ステミン」(STEM CELL INDUCING FACTOR, 幹細胞[ステムセル]誘導因子: 略してSTEMIN)と名付けました。

ステミン遺伝子は、切断した葉で幹細胞に変化しようとしている細胞で働いていることがわかりました。また、ステミン遺伝子とそれに近縁な2つの遺伝子の計3遺伝子を壊すと、切断した葉での幹細胞への変化が遅れることがわかりました。つまりステミン遺伝子は、植物にもともと備わっている幹細胞を作り出すシステムで働いていることが明らかになりました。
ステミン遺伝子は何をしているのか?
ではステミン遺伝子は、どのようにして分化細胞を幹細胞へと変化させるのでしょうか? 分化細胞がそれぞれ異なる性質や機能を維持できるのは、それぞれの細胞特有の遺伝子だけを働かせる仕組みがあるからです。遺伝子の実体であるDNAは、ヒストンH2A、H2B、H3、H4の4種類からなるコアヒストンと呼ばれるタンパク質複合体に巻きつき、クロマチンと呼ばれている構造を作っています。なかでも、ヒストンH3の27番目にあるリジンがメチル化という化学修飾を受けると、そのまわりの遺伝子が働かないようにクロマチンが閉じた状態になります。この修飾によって、分化した細胞では不必要な遺伝子の働きが抑えられています。
ステミン遺伝子からできるタンパク質は、多くの遺伝子の働きを一括して調節する転写因子です。そこで私たちは、ステミン転写因子によって直接調節される遺伝子領域のヒストンH3の化学修飾状態を調べました。ステミン転写因子は、細胞分裂を促進する遺伝子を含む1,416個の遺伝子を直接調節していることわかりましたが、葉細胞では、これらの遺伝子領域のヒストンH3にメチル基が付加されており、遺伝子の働きが抑えられていました。ところがステミン転写因子が働き始めると、そのメチル化レベルが減少し幹細胞化に必要な多くの遺伝子が働き出すことがわかりました。
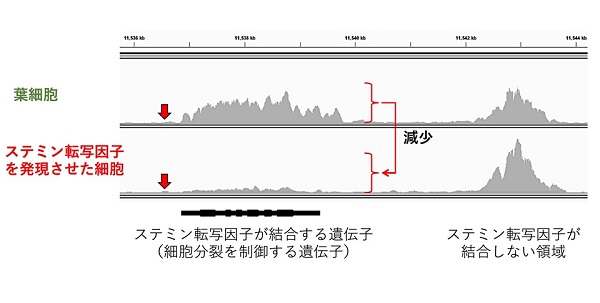
ステミン遺伝子からつくられたステミン転写因子は特定のDNA領域に結合すると、何らかの仕組みによってステミン結合領域近傍の遺伝子のヒストンH3K27me3修飾レベルが減少する。赤の矢印は、ステミン転写因子が結合する位置。
分化細胞を幹細胞へと変化させる仕組みが見えてきた
以上の結果から、分化細胞から幹細胞へと変化するときの過程が見えてきました。分化した葉細胞では、ヒストンH3の27番目のメチル化によって幹細胞化に必要な遺伝子群の働きが抑制されているので、葉細胞の性質を保つことができます。ところが葉が切断されると、切断面に面した葉細胞でステミン転写因子が蓄積し始め、特定のDNA配列にステミン転写因子が結合し、ヒストンH3のメチル化が外れます。その結果、これまで抑制されていた細胞分裂や細胞成長を促進させる遺伝子を含む多数の遺伝子が働き始め、それらの遺伝子の働きにより葉細胞が幹細胞へと変化するという仕組みが明らかになりました。
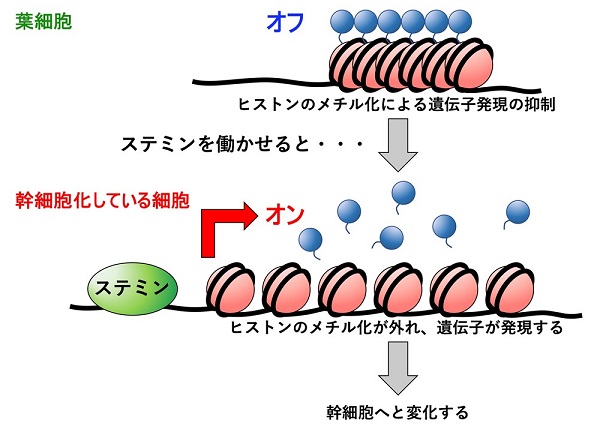
葉細胞では、幹細胞へ変化するときに働く多数の遺伝子がヒストンのメチル化によって転写が抑制されている(上)。ステミン遺伝子が発現すると、ステミン転写因子がその遺伝子領域に結合し、ヒストンのメチル化が外れて、幹細胞化に必要な遺伝子の発現が誘導される(下)。
転写因子はもともと多くの遺伝子の働きを一括して調節できますが、ステミン転写因子は、ヒストンの化学修飾によって厳格に働きが抑えられている幹細胞化に必要な遺伝子のヒストンH3のメチル化を外す働きと、メチル化レベルが減った遺伝子の発現を誘導させる2つの働きをしていると考えられます。
ステミン遺伝子は、イネやバラといった農作物や園芸植物などの植物にも存在しています。これらの植物におけるステミン遺伝子の機能はまだ不明ですが、コケ植物のようにステミン遺伝子の働きを調節することで簡単に増やせるようになるかもしれません。
一方で、ステミンがどのような仕組みでヒストンのメチル化レベルを減らすかについては未解明です。今後は、この点を明らかにすることで、植物の分化細胞を幹細胞に変える仕組みの全貌解明につなげていきたいと思っています。
参考文献
*Masaki Ishikawa, *Mio Morishita et al.(*共同筆頭著者)”Physcomitrella STEMIN transcription factor induces stem cell formation with epigenetic reprogramming” Nature Plants 5, 681-690 (2019)
DOI: 10.1038/s41477-019-0464-2 (https://doi.org/10.1038/s41477-019-0464-2)
この記事を書いた人

-
基礎生物学研究所 生物進化研究部門 助教
名古屋大学大学院修了後(博士(理))、ロックフェラー大学博士研究員、ERATO長谷部分化全能性進化プロジェクト博士研究員を経て、2012年より現職。専門は、植物幹細胞生物学と植物分子生物学。植物に特徴的な環境に応じた発生や分化を調節する仕組みについて、こつこつと研究しています。



