オタマジャクシの集団生活 – 成長で変わる付き合いのルール
いろいろな「群れ」
ヤマアカガエルのオタマジャクシには、他個体のサイズと血縁関係の両方を識別する能力があります。オタマジャクシは、発達に伴って群れる相手の好みも変化させます。ではなぜ、両生類の幼生にこのような性質があるのでしょうか。
私たちヒトを始め、動物の多くは群れを作って生活をしています。なぜかといえば、それは利益があるからです。その利益によってさまざまなタイプの「群れ」が存在します。捕食者から身を守るための群れ、探索効率を上げるための群れ、共同して繁殖するための群れ、複雑な社会をもつ群れ、などです。英語で「群れ」は、aggregation、swarm、schooling、flock、herd、groupと群れ方や動物の分類群に合わせいろいろな用語をもつのに対し、日本語はこれにうまく対応していません。
群れの在り方の違いは、利益とコストについての問題と捉え直すと、少し整理できそうです。群れも良いことだけでなく、個体レベルで損失(コスト)も発生します。第一に、病原菌に対する感染リスクの上昇です。群れることで個体間距離が小さくなりますし、集団として定住すれば衛生面の問題が生じます。次に、食料の奪い合いといった資源競争です。これらのコストは個体の適応度を下げます。集団生活の維持にはコストがかかります。このため、多くの動物の群れは一定しておらず、集合と離散を繰り返します。個体は常に利益とコストを天秤にかけて行動選択をしているのかもしれませんし、環境が安定していれば、迷う余地もなく単純なルールに従っているだけかもしれません。
オタマジャクシの集団生活
大学院時代にずっとヒキガエルを対象に遺伝的多様性や繁殖行動を研究してきた私は、毎春、池の中のオタマジャクシが頻繁に群れているのを観察しながら、彼らが何をしているのかずっと気になっていました。くっついたり離れたりといった個体間相互作用が起こっているのですが、各々がどう認識しているのか、ただの動く物と認識しているのか、群れるのは同種なのかどうか、また兄弟かどうか、他個体を識別しているのか、といった問題に興味を持ちました。
オタマジャクシの群れには保温効果や採餌効率の上昇、そして捕食者に対する防衛(希釈効果)といった利益があります。小さなうちは特に、単独でいるより群れでいる方が安全でしょう。群れを作る個体のあいだで利益とコストが同一ならば、誰と群れるのが最適か? などと考慮する必要もなさそうです。しかし、個体の発達に大きな差があればどうでしょうか。体のサイズや発達程度に差があれば、群れから得られる利益とコストにも違いが出ると予想されます。
発達段階の早い小さな個体にとって、発達が進んだ大きな個体は脅威になります。同じ時期に孵化した幼生の発生は基本的に同調するので、孵化時期が異なれば差が生じます。餌資源が限られていれば、集団内に競争が起こり、さらに個体間で発達に差がつきます。繁殖期に幅のある種では、遅生まれは早生まれの個体に捕食されるリスクに晒されます。また同じ他個体でも、兄弟か否かを識別するかもしれません。実際、いくつかの種では血縁個体への共食いを避ける傾向があります。遺伝子レベルで考えれば、血縁競争の上昇は好ましくないでしょう。
成長で変わる個体関係
集団内に競争がある場合、個体は群れる相手をどのように選択しているのでしょうか。血縁関係が影響しているか、あるいは発達差だけが影響しているのでしょうか。私はこの疑問に応えるのに、幼生間に発達差のあるヤマアカガエル(Rana ornativentris)は最適な種だと思いました。

海外の先行研究に両生類の幼生に血縁認識能力があるという報告が多く存在しています。血縁認識をするオタマジャクシは、匂いを頼りに兄弟と群れる傾向があるそうです。しかし血縁者は形態(サイズ)も似ています。これまでの研究では、血縁者のサイズまでは考慮していませんでした。
小さな個体は自分と同じサイズの個体と群れることを好むかもしれません。また、発達の進んだ大きな個体は、血縁関係にある小さな個体との競争を避けようと、血縁関係にない小さな個体と群れることを好むかもしれません。発達段階の異なる血縁個体を用意すれば、血縁関係とサイズ差を同時に検証できるはずです。
両生類の幼生の発達は水温や密度の影響を受けます。総研大(神奈川県葉山町)周辺に生息するヤマアカガエルの6つの卵塊を収集し、環境条件を調整することで発達段階の異なる2つのグループ(冷温に置いて発生を遅らせた小さな個体グループと、常温で通常時期に発生させた大きな個体グループ)を用意しました。この2つのグループを用いて、群れる相手の好み(社会的選好性)を調べる選択テストを行いました。

サイズと血縁関係、双方を認識
選択テストでは水槽を3つに区切り,中心部に観察対象である試験個体(大きな個体、または小さな個体)を入れ、水槽の左右に2匹の刺激個体を配置しました。刺激個体の組み合わせは、サイズ(大きな個体、または小さな個体)と、試験個体との血縁関係(同じ卵塊から生まれた血縁個体か、別の卵塊から生まれた非血縁個体か)から、4つのパターンで実験を行いました。
実験では、中心部に置かれた観察対象である試験個体が、どちら側により長く滞在するか、試験個体の動きを80分間撮影し、トラッキングソフトを使いそれぞれの滞在時間を算出しました。この滞在時間を比較し、解析を行った結果、大小どちらの試験個体も、小さな刺激個体を選ぶ時間の方が長く、大きな刺激個体を避ける傾向があることがわかりました。
また、血縁関係を考慮すると、大きな試験個体と小さな試験個体のあいだで、違いがありました。小さな試験個体では相手との血縁関係に関係なく大きな刺激個体よりも、小さな刺激個体を好みました。一方、刺激個体が両方とも非血縁である場合、大きな試験個体は、大きな刺激個体よりも小さな刺激個体を好みました。このような結果は、小さな刺激個体が血縁個体である場合にはみられませんでした。このことから、大きな個体は、血縁関係にある小さな個体と群れる傾向を持たないと言えます。
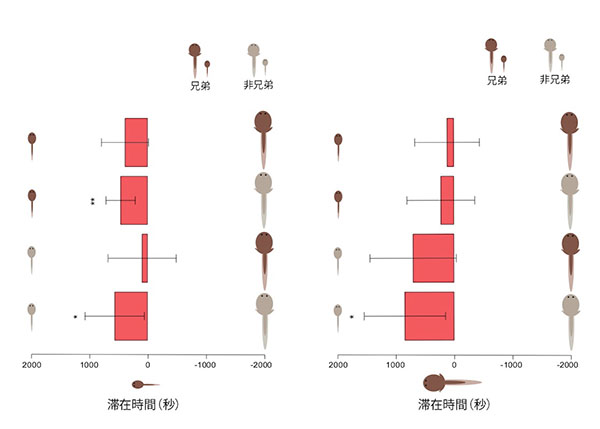
(a)小さな試験個体のテスト結果(上から30、28、24、24回試行)。統計的に有意だったテストパターンは、刺激個体が小さな兄弟と大きな非兄弟の時(* P<0.05, 対応のあるt検定)と、大小ともに非兄弟の時(** P<0.01, 対応のあるt検定)である。
(b)大きな試験個体のテスト結果(上から23、22、21、20回試行)。統計的に有意だったテストパターンは、大小の刺激個体が共に非兄弟の時のみ(** P<0.01, 対応のあるt検定)である。
仲間であることと集団の維持
ヤマアカガエルの幼生は、相手の大きさと血縁関係の両方を同時に考慮していることがわかりました。小さな個体が血縁の有無にかかわりなく小さな個体を好むのは、群れることに利益があるからでしょう。大きな個体同士では群れないのは、競争を避けるためと考えられます。大きな個体が小さな血縁個体を好まない理由についてははっきりとはわかりませんが、結果として血縁者間での競争緩和に繋がっているでしょう。どのようにして相手のサイズや血縁関係を認識しているのかについてもわかっていませんが、ヤマアカガエルの幼生の持つ「群れる相手の好みにおける発達の変化」は、集団内の競争の調整に効果があると考えられます。
サイズと血縁関係が両方関わる社会的選好性の報告はこれまでに例がなく、本発見によって幼生が群れを作る際のルールが世界で初めて明らかになりました。オタマジャクシが相手のサイズや血縁関係を識別できるというのは驚きですが、このような性質を持つことは集団の維持に役立っているのでしょう。しかしこの性質が変態後にどのような影響を与えるかについては、まだわかっていません。
なぜ多くの動物が群れを作るのか、どのように集団生活を維持しているのか、それぞれの種の生活史や進化的背景を考慮して研究を進めていくことで、全体像が明らかになっていくと考えています。
参考文献
Hase, K. & Kutsukake, N. 2019. Developmental effects on behavioural response for social preferences in frog tadpoles, Rana ornativentris. Animal Behaviour 154:7–16
この記事を書いた人

- 総合研究大学院大学先導科学研究科・特別研究員として、両生類を対象に集団内の多様性と行動生態について研究しています。特に、血縁認識能が社会性の進化に与えた影響について興味を持っています。客観的な視点で材料を選んできたつもりですが、調べているうちに好きになってしまう癖があります。ヒキガエルが大好きです。ウシガエルを料理できます。
この投稿者の最近の記事
 研究成果2019年9月26日オタマジャクシの集団生活 – 成長で変わる付き合いのルール
研究成果2019年9月26日オタマジャクシの集団生活 – 成長で変わる付き合いのルール


