鳥の糞から柑橘類の葉へ – アゲハ幼虫の変身を制御する遺伝子の発見
アゲハチョウ幼虫の紋様は変化する
アゲハチョウは私たちの身近で見られる蝶です。日本では、ナミアゲハ以外にもナガサキアゲハやクロアゲハなど10種類以上のアゲハチョウが知られていますが、その幼虫の多くは、若齢時は白と黒の紋様からなり、鳥の糞に擬態しているといわれています。一方、これらの幼虫は最終齢になると、食草である柑橘系の葉に紛れるような緑色に変わります。

鳥の糞への擬態と柑橘系の葉へのカムフラージュにより、捕食者である鳥から捕食されにくくなると考えられています。しかし、どのようにして2つの擬態紋様が生じ、また切り替わるのか、その分子機構はほとんどわかっていませんでした。今回、私たちの研究グループはナミアゲハの幼虫の紋様を切り替える遺伝子とその作用メカニズムの一端を明らかにしました。
幼虫紋様の変化を制御する遺伝子の探索
まず、2つの擬態紋様の切り替えを決定する時期(4齢幼虫の初期:4齢脱皮後0時間、6時間、12時間後)に3つの紋様領域(鳥の糞模様の黒、鳥の糞模様の白、5齢幼虫の目玉模様)のRNAを調製し、RNA sequencing法によりそれぞれの領域や時期で発現しているRNAを比較しました。その結果、黒、白、目玉領域で特異的に発現している遺伝子が複数(合計で20個)同定されました。
そのなかで、Clawless (Cll)(目玉模様特異的)、abdominal-A (abd-A)(黒・白領域特異的)、Abdominal-B (Abd-B)(黒領域特異的)といったホメオボックス遺伝子は、4齢初期の特定の時期に、括弧内で示した特定の紋様で発現変動していることがわかりました。4齢初期から3日ほどすると脱皮期(5齢脱皮期)になりますが、このときに4齢幼虫のクチクラが脱ぎ捨てられ、新たに5齢幼虫の擬態紋様が描かれたクチクラが形成されます。
3つのホメオボックス遺伝子はプレパターン期にのみ幼虫紋様を変化させうる
アゲハチョウのような鱗翅目昆虫の多くは、一般的にRNAi(RNA干渉法)が起こりにくく、遺伝子の機能を調べることが困難でした。数年前、私たちのグループではエレクトロポレーション法を利用して遺伝子機能を調べることに成功し、今ではさまざまな発生ステージのアゲハチョウで遺伝子の機能を調べることができるようになりました。
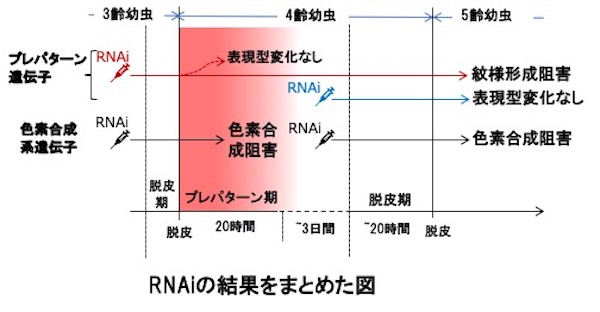
そこで、エレクトロポレーション法で3つのホメオボックス遺伝子に対するsiRNAを3齢幼虫時にそれぞれの紋様領域に導入し、RNAiにより遺伝子発現を抑制しました。そうすると4齢幼虫時(鳥の糞型紋様)の紋様は変化しなかったのに対し、5齢幼虫時の紋様形成は阻害されました。図はabd-Aの機能を阻害した結果ですが、5齢幼虫の紋様の一部が消えています。
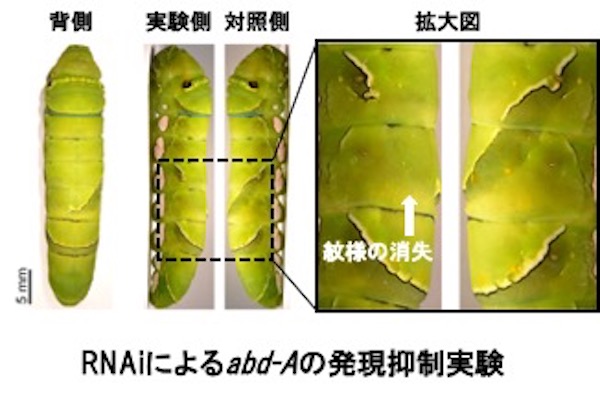
しかし、4齢初期の紋様の切り替え決定時期を過ぎてからsiRNAを注入して同様の処理をしても、5齢幼虫の紋様形成は阻害されませんでした。一方、メラニンなどの着色に関与する遺伝子に対して同様の処理をすると、4齢幼虫も、5齢幼虫も紋様の一部が形成されなくなりました。これらの結果から、上記のホメオボックス遺伝子は紋様のプレパターンを4齢の脱皮期(5齢幼虫の紋様を作る)よりも前の、紋様切り替え時期(プレパターン期)に決定していることが明らかになりました。
幼若ホルモンが幼虫紋様を変化させる
アゲハ幼虫の鳥の糞型から柑橘葉への擬態紋様の切り替えは、体内の幼若ホルモンJHの低下によって決定されることが以前からわかっていました。4齢初期にJHを塗布して体内のJH濃度をあげると、5齢幼虫になっても鳥の糞のままで紋様が切り替わりません。そこで、このJH処理を施した幼虫で上記のホメオボックス遺伝子の発現を調べると、処理していない幼虫に比べて、低下していました。この結果は、JH濃度の低下により(つまりJH濃度が低下するプレパターン期に)、それぞれのホメオボックス遺伝子の発現が誘導されることを示唆しています。
以上の結果から、紋様の切替え(5齢幼虫も紋様のプレパターン)は、実際に紋様が形成される脱皮期よりかなり前に、JHの低下によって誘導されるホメオボックス遺伝子などを介して決定されることがわかりました。擬態の複数の紋様がいくつかのホメオボックス遺伝子により事前に決められていることや、ホルモンによってこれらの遺伝子の発現が制御されていることなどが初めて明らかになりました。
日本には生息しないアゲハチョウの仲間には、蛹になるまで鳥の糞型の幼虫のままである種が知られており、上記のような紋様の切替えしくみは祖先的なアゲハから進化の過程で獲得されたと考えられます。また、セリ科の植物を食べるキアゲハの幼虫のように柑橘系の葉と食べるアゲハとはまったく異なる終齢幼虫のパターンがどのように切り替えられるのかにも興味がもたれます。
今回得られた結果は、他の擬態における紋様形成におけるメカニズムは進化を知るうえでも、大きな手がかりを与えると考えられます。また、今後はまだ確認をしていないホメオボックス以外の候補遺伝子の機能を知ることで、擬態紋様の形成メカニズムやその進化プロセスをより詳細に知ることができるようになると期待しています。
参考文献
- Jin. H., Seki, T., Yamaguchi, J., Fujiwara H. (2019) Prepatterning of Papilio xuthus caterpillar camouflage is controlled by three homeobox genes: clawless, abdominal-A, and Abdominal-B. Sci. Adv., 10, eaav7569.
- Futahashi, R., Shirataki, H., Narita, T., Mita, K., Fujiwara, H. (2012) Comprehensive microarray-based analysis for stage-specific larval camouflage pattern-associated genes in the swallowtail butterfly, Papilio xuthus. BMC Biol. 10, 46.
- Futahashi, R., Fujiwara. H. (2008) Juvenile hormone regulates butterfly larval pattern switches. Science 319, 1061.
この記事を書いた人

- 東京大学大学院・新領域創成科学研究科・先端生命科学専攻・教授。理学博士。1957年神戸市生まれ。東京大学・大学院理学系研究科・生物化学専攻修了後、国立予防衛生研究所(現感染研)・研究員、東京大学・理学部・動物学教室・講師、ワシントン大学・動物学部・研究員、東京大学・大学院理学系研究科・生物科学専攻・助教授、同大学院・新領域創成科学研究科・准教授などを経て現職。昆虫を主な研究対象として、擬態や変態の分子機構、テロメアや利己的遺伝因子の進化などの解明に分子生物学的な切り口から取り組んでいる。



