酵母の巧妙なフェロモン認識システム – 曖昧さが進化の鍵!?
動物から微生物まで多くの生物では、「性フェロモン」という物質を使って、異性に存在を知らせ交配しています。そのため、何らかの原因によりフェロモンもしくはその受容体タンパク質が遺伝的に変化すると、これらの変化した個体は交配相手を見つけられなくなり、結果として集団から隔離されてしまいます。
このようなリスクを抱えながら、どのようにしてフェロモンの多様性は生まれるのでしょうか? ここでは、私たちのこれまでの酵母の研究を踏まえて、その仕組みを紹介したいと思います。
酵母の性フェロモン
酵母にも動物と同じように2つの性があります。私たちが研究対象にしている分裂酵母(Schizosaccharomyces pombe)の2つの性は、Plus型(P型)とMinus型(M型)と呼ばれています。
動物では雄と雌で外観が異なるのが普通ですが、分裂酵母の場合は見ただけでは性別はわかりません。しかし、分裂酵母はP型とM型の細胞のあいだでしか交配(=細胞融合)はしないのです。では、どうやって分裂酵母は異性を識別しているのでしょうか?
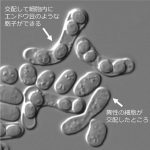
それは、それぞれの細胞が「性フェロモン」を分泌して、異性を刺激して交配へと導くからです。酵母のフェロモンはアミノ酸がつながったペプチドであり、これが異性細胞の細胞膜にある受容体に結合します。するとフェロモンを受容した細胞は、「異性が近くにいる!」と張り切って交配相手を探すようになります。
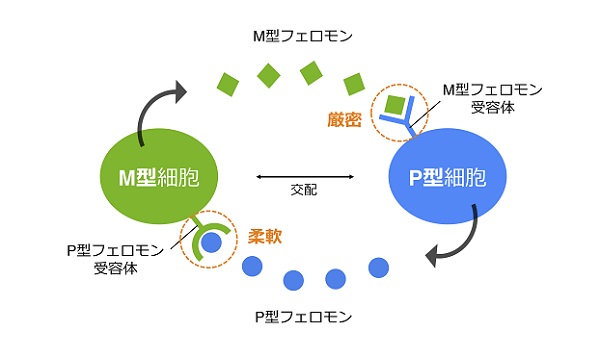
フェロモンの構造が少しでも変わると、受容体とは結合できなくなるので、フェロモンと受容体はちょうど鍵と鍵穴のような関係になっています。そのため、フェロモンの構造が異なる酵母とは交配できません。この「厳密性」が同種間での交配を確実にしており、他種からの生殖隔離(2つの個体群のあいだで生殖が行われない状態)に役立っています。
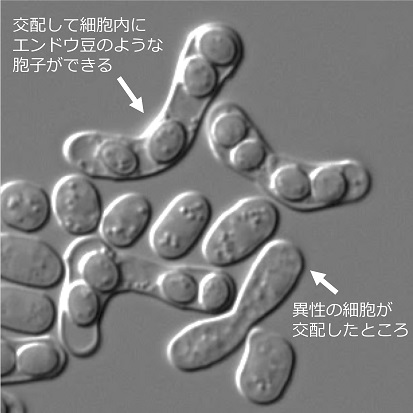
異性細胞間でフェロモンをやりとりすることにより交配する。細胞融合すると胞子(=子孫)が4つ作られる。
“新しい種”のように振舞う人工的な酵母
では、フェロモンを無理やり変えてやるとどうでしょうか? 受容体に結合できなくなり、交配ができなくなるはずです。分裂酵母では、フェロモンの構造は直接遺伝子によって決められています。そこで、遺伝子操作がしやすいという酵母の利点を生かして、私たちはフェロモンの遺伝子を自由自在に変えることにしました。
注目したのは、9つのアミノ酸からなるM型フェロモンです(ちなみにP型フェロモンは23つのアミノ酸です)。どうせなら網羅的に変えてやろうと考え、構成するアミノ酸のひとつを別のアミノ酸に順々に置き換え、合計152種もの変異型フェロモンを作りました。こうした異なる構造のフェロモンが受容体に正しく結合できるかを調べたところ、そのうち35種がまったく機能しませんでした。
さて、ここからがポイントです。今度は受容体の構造を変えてやれば、この構造が変わったフェロモンを受容できるのではないかと考えました。つまり、形の変わった鍵に合わせて、鍵穴も変化させることができれば偶然ぴったり合うのではないかと閃いたのです。
この戦略は単純なように思われましたが、想像以上に苦労しました。2年以上に及ぶ努力の末に、およそ65万以上の受容体候補のなかからようやく目的の変異型受容体を見つけることができました。
この変化したフェロモン/受容体のペアを持つ酵母は、きちんと交配して子孫を残すことができますが、元の正常な酵母とはまったく交配しません。こうして、私たちはフェロモンと受容体を人工的に改変することによって、野生型から生殖隔離された「新しい酵母」を創ることに成功したのです。
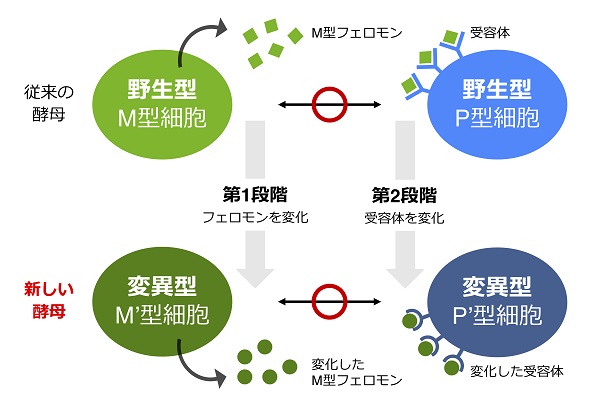
フェロモンとその受容体遺伝子を変化させることにより、従来の酵母とは交配しない「新しい酵母」を人工的に創った。
自然界で2つのフェロモンが非対称に変化する不思議
しかし、ここで疑問が生じます。自然界では、どのようにしてフェロモンとその受容体の組み合わせが変化するのでしょうか? 私たちはその答えを探るべく、世界各地に棲む野生の分裂酵母(S. pombe)を調査することにしました。
日本産の3株を含む、22カ国以上の国から単離された合計150株の分裂酵母を対象に、フェロモンと受容体の遺伝子を解析しました。
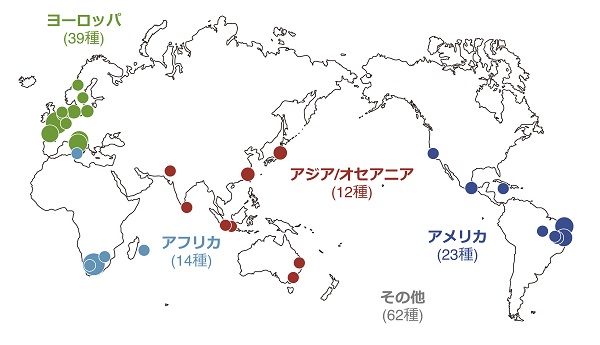
するとおもしろいことに、M型フェロモンとその受容体は完全に構造が保存されていたのに対し、P型フェロモンとその受容体は大きく変化していたのです。同じ種でありながら、自然界にはP型フェロモンは6種類、そしてその受容体は5種類もの多型が存在していました。
この2つのフェロモンに見られる多様性の違いは、分裂酵母(S. pombe)の近縁種であるSchizosaccharomyces octosporusにおいてもみられる普遍的な現象でした。つまり、分裂酵母では、M型フェロモン/受容体のペアは厳密に保たれていますが、P型フェロモン/受容体のペアは比較的柔軟性に富んでおり、その認識も曖昧になっているようです。
P型フェロモンは近縁種にも作用可能
次に私たちは、分裂酵母に自身のフェロモンの代わりに、他種のフェロモンを作らせるとどうなるかに興味を持ち、早速調べることにしました。
そこで、分裂酵母(S. pombe)と近縁種(S. octosporus)のあいだでフェロモンをそっくりそのまま入れ替えてやると、なんとP型フェロモンは他種のものと交換しても機能したのです。つまり、S. pombeのM型細胞はS. octosporusのP型フェロモンを分泌するP型細胞と交配することができ、逆もまた然りでした。
一方で、M型フェロモンは他種のものと交換することはできませんでした。つまり、M型フェロモンは種特異的に作用し、一方P型フェロモンは種を超えて作用するということです。実際にペプチドを化学合成して細胞に処理すると、M型細胞は他種のP型フェロモンに反応する様子が見られます。しかし、酵母の交配にはM型フェロモンとP型フェロモンによる「相互の」フェロモン認識が必須なため、通常、間違って異なる種と交配することはないのです。
酵母が“非対称な”フェロモン認識システムを持つ理由
今回私たちは、分裂酵母のM型フェロモン/受容体のペアは厳密に保たれている一方で、P型フェロモン/受容体のペアは柔軟に変化することができるという「非対称性」を発見することができました。
これらのフェロモン認識における「厳密さ」と「柔軟さ」の両方を兼ね備えた認識システムは、酵母が同種間の交配を保ちつつも、フェロモンの多様性を生み出すことを可能にしていると考えられます。こうして、変化したフェロモンを特異的に結合できるように変化した受容体が集団内に出現すると、それらの子孫は元々の集団から隔離され、新たな種として進化していくのかもしれません。
これまで、交配という重要なイベントを司るフェロモンの認識は厳密であると考えられてきましたが、同時に「曖昧さ」も備わっており、これが生物の進化の原動力になっていると考えられます。今後は、片方の「鍵と鍵穴」の認識を(わざと?)緩くしておくことに、どのような生物学的なメリットがあるのかを解明していきたいと思っています。
参考文献
この記事を書いた人

-
清家 泰介(Seike Taisuke)
理化学研究所 生命機能科学研究センター・基礎科学特別研究員。
2014年、大阪市立大学 大学院理学研究科・後期博士課程修了 (日本学術振興会特別研究員DC1)、博士 (理学)。その後、国立研究開発法人 産業技術総合研究所・博士研究員 (2014年~2015年)、情報・システム研究機構 国立遺伝学研究所・日本学術振興会特別研究員PD (2015年~2018年)を経て、2018年より現職。分裂酵母を使って、フェロモン/受容体システムによる生殖隔離・種分化のメカニズムを研究中。
この投稿者の最近の記事
 研究成果2019年3月11日酵母の巧妙なフェロモン認識システム – 曖昧さが進化の鍵!?
研究成果2019年3月11日酵母の巧妙なフェロモン認識システム – 曖昧さが進化の鍵!?


