被子植物が2組のカップルを正確につくるための、男女のやりとり – 「重複受精」の新たな因子を探る
人の生活と重複受精との関わり
私たちが日ごろ食べている野菜や果物、穀物、そして目を楽しませてくれる花々は、どれも生活にとても身近な植物です。これらの大部分は、植物のなかでも「被子植物」に分類されます。被子植物はその大きな特徴として、「重複受精」と呼ばれる独特の生殖方法を進化の過程で獲得しています。重複受精の結果、被子植物は種子を形成し、次の世代の植物を誕生させます。
被子植物では「花粉」がオスの組織に、「胚珠」(将来種になる組織で、雌しべの中に形成される)がメスの組織に該当します。花粉の中にはオスの生殖細胞である2つの「精細胞」が、胚珠の中には2種類のメスの生殖細胞として「卵細胞」と「中央細胞」が形成されます。
受粉後、花粉は花粉管を発芽して伸長しながら、2つの精細胞を胚珠まで運びます。胚珠の中まで伸びてきた花粉管は先端を破裂させて、精細胞を放出します。その結果、2つの精細胞、卵細胞と中央細胞の計4つの細胞が対面します。やがて、精細胞のひとつは卵細胞と、もうひとつは中央細胞と融合して、受精が起こります。
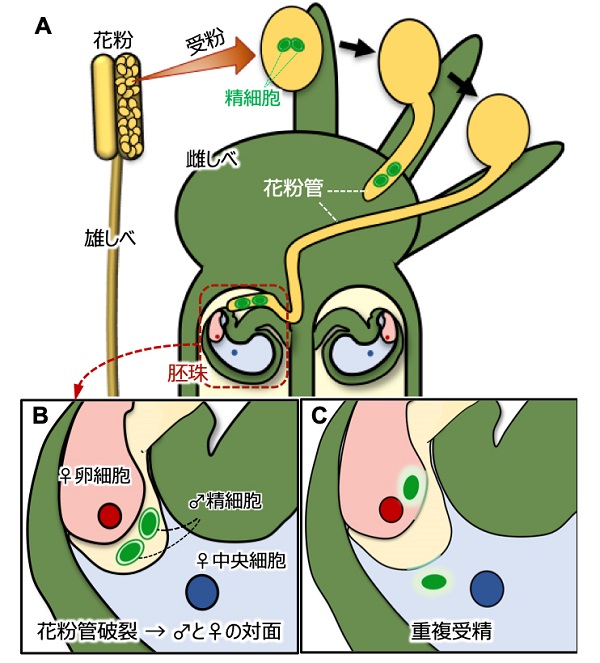
(A) 花粉が雌しべの頭に受粉すると、花粉管が発芽・伸長して2つの精細胞を胚珠まで運ぶ
(B) 花粉管が破裂して2つの精細胞が卵細胞および中央細胞の間に送り込まれる
(C) 精細胞のひとつは卵細胞と、もうひとつは中央細胞と細胞膜融合を介して受精する(重複受精)
このように、被子植物の生殖プロセスでは2種類のオス・メスのカップル間での受精がほぼ同時に起こるので、「重複受精」と呼ばれています。受精した卵細胞は胚(次世代の植物)に、中央細胞は胚乳(胚の養育組織、お米でいうと可食部に相当)に発達し、ひとつの種子を完成させます。重複受精のうちどちらかが失敗してしまうと、種子は得られません。
さて、現在お店で売られている植物のほとんどは、人間が意図的に改良、すなわち育種を重ねてきた賜物でもあります。植物を改良する主な方法に、交雑育種法があります。異なる特徴(=遺伝子)を持つ両親同士を掛け合わせ、雑種を得る方法です。この方法は、植物の重複受精現象を利用しています。
今でこそ植物での交雑育種は当たり前の概念ですが、実は、人類が学術的に植物にも性があることを知ったのは17世紀の終わりごろです。動物と同じように植物にもオスとメスがあることを知ってから、人は意思を持って交雑育種による植物改良を進め、現在に続いています。
重複受精研究の歴史
植物の性の発見から約200年後の1898年に、ロシアの植物学者ナワシンによって重複受精現象は発見されました。その後、さまざまな種で重複受精の観察研究が行われてきましたが、最近では遺伝子解析用のモデル植物でもあるシロイヌナズナで詳しい仕組みが明らかになり始めています。
オスとメスの生殖細胞が出会ってから受精に至るまでには、互いの「認識」と「接着」の後に、「融合」するプロセスを経ることがわかってきました。そして、これらの各ステップを司るタンパク質が発見されたのは21世紀に入ってからです。
2006年に初めての重複受精因子としてオスのGCS1が発見され、メス生殖細胞との融合に関わることがわかりました。続いて2014年にオス因子であるGEX2がメス生殖細胞との接着に関わることが報告されました。
これまでに、シロイヌナズナの2つの精細胞はまったく同一の双子であり、どちらも卵細胞または中央細胞と受精できることがわかっています。そして、被子植物は正確に、「精細胞と卵細胞」&「精細胞と中央細胞」の2組の男女カップルを成立させて、それぞれを結婚させています。双子のオスが両方とも片方のメスと結婚してしまうことはありません。
正確な2組の結婚のためには、オスとメスのあいだで執り行われる「認識」ステップが深く関わっていると考えられますが、このステップに関与する因子はまだ同定されていませんでした。
2組のペアはどのように成立している?
重複受精はオスとメスの生殖細胞間のやりとりによって進行するので、そのやりとりを担うタンパク質は、各生殖細胞の最外層となる原形質膜に存在すると考えられます。受精因子としてはたらくタンパク質を新たに見つけるために、私たちは生殖細胞の膜タンパク質を回収し、その中から受精に関わるものを見つけ出すアプローチをとりました。
まず、オスの生殖細胞の回収法が確立されているテッポウユリをスタート材料として、膜タンパク質を精製し、そこに含まれるタンパク質の種類を網羅的に解析しました。このなかから、シロイヌナズナにも類似のタンパク質が存在するもの、かつ花粉で遺伝子発現が高いものを候補として絞り込みました。
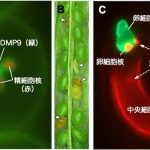
さらに、解析候補に挙がった遺伝子に蛍光タンパク質の遺伝子を融合させてシロイヌナズナに導入し、どこで蛍光が観察されるかを指標として、解析タンパク質の局在を調べました。その結果、「DMP9」というタンパク質が精細胞の原形質膜に特異的に局在することが示されました。
精細胞膜に局在するDMP9がはたして受精に関わるのか? この謎を調べるために、DMP9の遺伝子の発現を人工的に抑制したシロイヌナズナ遺伝子組換え体 DMP9KDを作りました。すると、このDMP9KDを父親(花粉親)に用いたときにだけ、発達が途中で停止してしまった異常な種子が20%弱観察されました。さらに重複受精を観察してみると、DMP9KDの精細胞のうちひとつだけが受精できずに残されていました。
この受精異常パターンについて、データ数を重ね統計解析した結果、DMP9KD精細胞は有意に卵細胞と受精できなくなる、あるいは卵細胞との受精が遅延することが明らかになりました。その結果、種子の発達が途中で停止してしまっていたのです。こうして、DMP9が新たな受精因子であると決定付けられました。
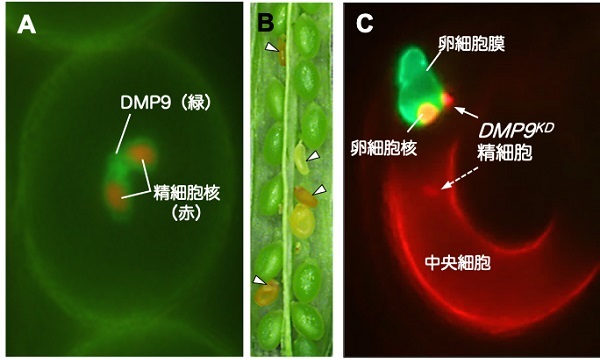
(A) DMP9は精細胞の細胞膜に局在する
(B) DMP9KD遺伝子組換え体では▽で示すように、途中で発達が停止する種子が20%弱観察された
(C) 重複受精において、DMP9KD精細胞は中央細胞とは受精できるが(点線矢印)、卵細胞とは受精できない(実線矢印)
今回の解析では、DMP9が減っている双子のオスに対して、2種類のメスは異なる対応をしたことになります。ちなみに、卵細胞と受精できなかったDMP9KD精細胞は、卵細胞に接着まではしていました。過去の研究で、メスの生殖細胞まで精細胞が3つ以上運ばれてくる異常が起きた場合、中央細胞は2つ以上の精細胞と受精してしまった、という報告があります。
これらの結果を合わせて考えると、卵細胞はひとつの精細胞とだけ接着したことを、DMP9を介して確認しているのではないか、と推測されます。
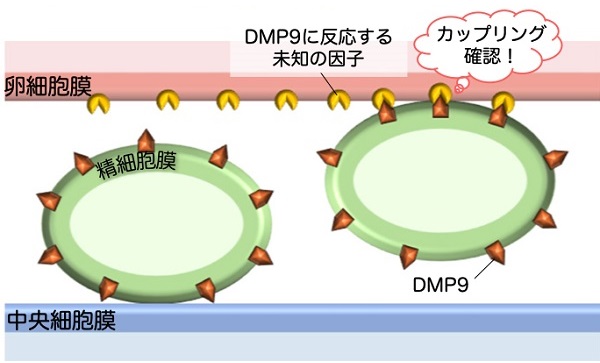
卵細胞膜上にはDMP9に反応する未知の因子が存在し、ひとつの精細胞と接着したときにカップリングを確認する。中央細胞にはDMP9と反応する因子がなく(あるいは少なく)、別の仕組みによるカップリング認識機構があると考えられる。
通常は2つやってくる精細胞に対し、卵細胞がひとつとだけ受精しようとすれば、残った精細胞が必然的に中央細胞と受精するようになっているのかもしれません。今は仮説の段階ですが、この検証をするために、引き続きDMP9の分子機構の解明に取り組んでいます。
先に紹介したGCS1やGEX2も含め、被子植物で発見されている受精因子はすべてオス側の因子であり、これらと相互作用するメス側の膜タンパク質はまだ発見されていません。花の雌しべの中の胚珠内で起こる重複受精の謎のヴェールはまだ幾重にも重なっています。
しかしいつか、人類が当たり前のように利用してきた重複受精現象の全貌が徐々に明らかになれば、その知見を活かした新たな育種法の開発にも繋がるでしょう。それは次の世代の人たちの仕事になるかもしれませんが、今はヴェールを一枚一枚地道にはがしていき、その知見を後世に残していく努力を続けたいと思います。
参考文献
Takahashi T, Mori T, Ueda K, Yamada L, Nagahara S, Higashiyama T, Sawada H, Igawa T. (2018) “The male gamete membrane protein DMP9/DAU2 is required for double fertilization in flowering plants” Development, 145: dev170076
この記事を書いた人
- 千葉大学大学院園芸学研究科・植物細胞工学研究室・准教授。日本女子大学理学部物質生物科学科を卒業後、千葉大学大学院自然科学研究科に進学。植物バイオテクノロジー研究にて博士号(農学)を取得。植物改良に関する研究に携わるなかで、交雑育種の基盤現象である有性生殖の仕組みを把握することが重要と考え、近年は重複受精分子メカニズムの解明研究に取り組んでいる。