「陸のプランクトン」トビムシに秘められたオメガ3脂肪酸合成経路の発見
トビムシは、その名のとおり飛び跳ねる行動が特徴的な、翅をもたない小さな節足動物です。トビムシは六脚類(Hexapoda)に属しますが昆虫類(Insecta)とは異なる分類群であり、六脚類の中で昆虫類よりも早期に分岐した系統群のひとつです。自然界では、トビムシは主に表層土壌や落葉層に高密度に分布し、個体数は1平方メートル当たり数万匹に達します。トビムシは、陸上の食物網において重要な役割を担っており、菌類や枯死植物など幅広い有機物を摂食・分解するとともに、肉食動物の重要な栄養源となっています。本記事では、陸上食物網を支える小さなトビムシに秘められた、機能性脂質・オメガ3脂肪酸の産生能力についてご紹介します。

トビムシはオメガ3脂肪酸のEPAを生合成する
従属栄養動物は、自身の発育や生存にとって必要な有機化合物をすべては自己合成せず、特定の分子グループについては捕食する生物に依存します。必須アミノ酸やビタミン類がその最たる例ですが、脂質のカテゴリーではオメガ3脂肪酸が代表例として挙げられます。オメガ3脂肪酸のうち、エイコサペンタエン酸(EPA)などの、炭素鎖長が20以上の「長鎖オメガ3脂肪酸」(注釈)は、ヒトをはじめとした脊椎動物において免疫機能や循環器系の調節など多様な機能を担っていますが、脊椎動物自身は長鎖オメガ3脂肪酸を自己合成せず、食物から摂取しています。海や川の中では、微細藻類などのさまざまな水生プランクトンが長鎖オメガ3脂肪酸を生産し、これが食物連鎖を通じて魚類やエビ・カニ類などの消費者に利用・蓄積され、最終的にはヒトがこれらの水産物の恩恵に預かり長鎖オメガ3脂肪酸を取り入れています。
長鎖オメガ3脂肪酸を合成できるか・できないかは、その生物が保有する「脂肪酸不飽和化酵素」によって決まります。脂肪酸不飽和化酵素とは、脂肪酸に二重結合を導入する酵素のことであり、メチル基末端側に二重結合を導入する「methyl-endタイプの不飽和化酵素」と、カルボキシル基末端側に二重結合を導入する「front-endタイプの不飽和化酵素」が長鎖オメガ3脂肪酸の合成に必須です。長鎖オメガ3脂肪酸を産生する水生プランクトンはこれら2タイプの酵素を有しています。一方、陸上生物の多くは、これら2種類の酵素グループのどちらか、または両方を失っており、長鎖オメガ3脂肪酸の合成能が見出されている生物種は一部の線虫類などごく少数にとどまっています。脊椎動物の場合、methyl-endタイプの不飽和化酵素を失っています。また、陸上食物網の生産者である維管束植物や、分解者である菌類の場合、front-endタイプの不飽和化酵素を持たず長鎖オメガ3脂肪酸の供給源にはなりません。また、陸上食物網の低次消費者である昆虫類は、methyl-endタイプとfront-endタイプのどちらも欠いています。そのため、水圏から隔離された陸上食物網では、長鎖オメガ3脂肪酸が不足しがちな栄養素となります。
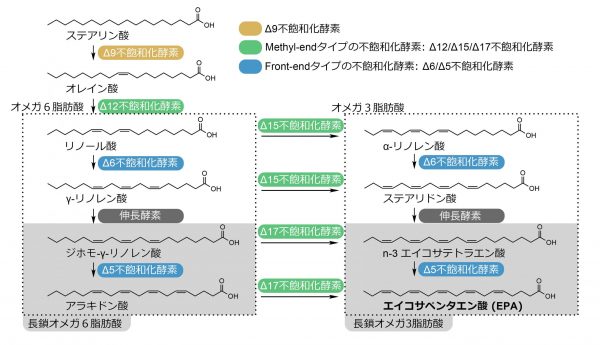
一方、前述のトビムシには、陸上生物のなかでは珍しく長鎖オメガ3脂肪酸のEPAが豊富に含まれます。たとえば、シロアヤトビムシという種では、総脂肪酸のうち5〜10%がEPAであり、湿重量1gあたりEPAは10mgほど含まれます。これは魚類に匹敵するEPAの含有量であり、トビムシは陸上生態系においてEPA生産者としての役割を担っていることが示唆されます。しかし、トビムシ類のEPA合成経路を担う酵素は明らかになっていませんでした。そこで私たちは、「トビムシ類はmethyl-endタイプとfront-endタイプの不飽和化酵素を保有している」という仮説を立て、シロアヤトビムシを含む4種のトビムシを対象に、EPA合成経路の解明に挑みました。
注釈:長鎖オメガ3脂肪酸とは、二重結合を複数有する多価不飽和脂肪酸のうち、メチル末端から数えて3および4番目の炭素に二重結合をもち、かつ炭素数が20以上の脂肪酸の総称です。
トビムシが持つ「非古典的なEPA合成酵素」の発見
まず、ガスクロマトグラフィー質量分析(GC-MS)を用い、トビムシ類においてEPAが主要な脂肪酸のひとつであることを確認するとともに、同位体標識した脂肪酸を用いた代謝活性解析により、トビムシ体内で確かにEPA合成が起こっていることを確認しました。次に、不飽和化酵素の候補を選抜するために、4種のトビムシを対象に、次世代シーケンシングによる遺伝子配列情報の取得と解析を行いました。その結果、front-endタイプの不飽和化酵素に類似した酵素をコードする遺伝子がすべての種において複数見出されました。しかし、methyl-endタイプの不飽和化酵素に類似する配列をコードする遺伝子は対象としたトビムシから検出されませんでした。この時点で当初描いていた仮説が崩れ、古典的なmethyl-endタイプの不飽和化酵素に依存しないEPA合成経路がトビムシに秘められている可能性が出てきました。
ひとまず、front-endタイプの不飽和化酵素に類似した酵素の活性を調べてみることにしました。対象とする遺伝子をクローニングし出芽酵母に発現させ、その酵母を特定の脂肪酸を添加した培地で培養し、脂肪酸のどの位置に二重結合が導入されるのかGC-MSにより解析しました。この基質変換実験により、リノール酸からアラキドン酸に至るまでのオメガ6脂肪酸の不飽和化を触媒する酵素群を同定することができました。では、アラキドン酸(オメガ6脂肪酸)からEPA(オメガ3脂肪酸)への変換を触媒する酵素はどのようなものなのでしょうか?いろいろと可能性を模索した末、ひとつの仮説として、「front-endタイプの不飽和化酵素に分類される酵素の一部はmethyl-endタイプの不飽和化酵素としての活性を有している」という考えに至りました。ここまで、タンパク質の一次配列の類似度をもとに酵素活性を推察していましたが、ときにタンパク質は一次配列からは予想されない機能を示すこともあります。この仮説を検証するために、アラキドン酸を基質として培地に加え、同様の基質変換試験を行いました。その結果、めでたく目的の新規酵素を見出すことができました。同定した「非古典的なmethyl-endタイプの不飽和化酵素」は、配列的にはfront-endタイプの不飽和化酵素に分類されますが、アラキドン酸からEPAへの変換を選択的に触媒するmethyl-endタイプの酵素活性を持ちます。従来のfront-endタイプ酵素と異なり、なぜアラキドン酸のメチル末端側に二重結合を導入できるのか、その構造的基盤は現時点では不明ですが、基質に結合するアミノ酸のいくつかに変異があり、それがこの酵素の基質認識の変容に繋がっている可能性を考えています。
トビムシは微細藻類から獲得したEPA合成酵素遺伝子を独自に進化させた?
六脚類に近縁とされるミジンコ類は、機能的なmethyl-endタイプとfront-endタイプの不飽和化酵素の明確なオルソログを持ちません。このことから、トビムシ類はEPA合成に必要な酵素遺伝子を別の生物グループから獲得(すなわち水平伝播により獲得)したことが予想されます。この可能性を検証するために、分子系統樹解析を行った結果、トビムシがもつ「front-endタイプの不飽和化酵素」と「非古典的なmethyl-endタイプの不飽和化酵素」は、長鎖オメガ3脂肪酸を産生する微細藻類などの微生物がもつfront-endタイプの不飽和化酵素とアミノ酸配列が近縁であることがわかりました。以上の結果から、トビムシの祖先は長鎖オメガ3脂肪酸を産生する微生物からfront-endタイプの不飽和化酵素をコードする遺伝子を獲得し、その遺伝子が重複・機能分化することにより独自のEPA合成経路が構築された、というシナリオを提唱しました。
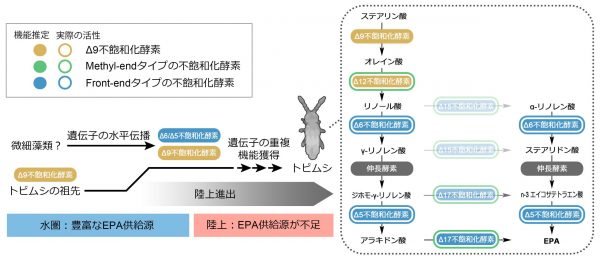
また、ここでは詳しく述べませんが、EPA合成経路の初期段階であるオレイン酸からリノール酸への変換に必要な、もうひとつのmethyl-endタイプの不飽和化酵素も同定しました。配列的にはステアリン酸からオレイン酸の合成を触媒する「Δ9不飽和化酵素」に類似する酵素群のうちひとつが、オレイン酸からリノール酸への変換を触媒していることがわかりました。Δ9不飽和化酵素は甲殻類や六脚類が普遍的に持つ酵素であるため、トビムシの進化の過程で内在的なΔ9不飽和化酵素が重複・機能分化したと考えられます。
トビムシは、六脚類のなかでは最初期に水圏から陸上へ進出したグループです。陸上という、EPAが少ない環境にトビムシたちが適応するには、微細藻類から獲得した遺伝子を進化させ、彼ら自身がEPAを生合成する必要があったと予想されます。そして、彼らが産生するEPAは、陸上食物網の上位に位置する脊椎動物のみならず、トビムシを捕食する肉食昆虫にとっても有益な栄養素となっている可能性もあります。もしそうなら、トビムシ自身や、トビムシを捕食する昆虫においてEPAはどのような生理機能を有しているのでしょうか。この点についても今後の研究で明らかにしていきたいと考えています。
おわりに -土や落葉にひそむ「翅のない虫」から見えてくるもの
前述のとおり、トビムシは落葉層や土壌表層を優占する小型節足動物であり、菌類や枯死植物から上位捕食者への物質循環の橋渡しを担う性質から「陸のプランクトン」と呼ばれています。今回の研究により、トビムシは脂肪酸合成の側面からも海のプランクトンと類似した性質を持っていることが明らかとなりました。
トビムシのほかにも、カマアシムシ、コムシなど、土壌や落葉にはさまざまな「翅のない六脚類」が息づいています。これらの六脚類は、陸上への適応や翅の獲得といった、昆虫が地上で大繁栄するに至った進化的変遷を研究するうえで重要な材料となります。トビムシ以外の翅のない六脚類もEPA合成経路を有しているのであれば、六脚類は祖先的にEPA合成能を有しており、有翅昆虫への進化の過程でその能力を放棄した可能性が考えられます。今後の研究では、六脚類基部に分類される系統群の脂肪酸合成経路とその進化的変遷や生理的意義を紐解き、脂肪酸クオリティの変容の観点から六脚類の進化を理解する研究に発展させていきたいと思っています。
参考文献(* 責任著者)
- Ohhara Y*, Sato A, Hirono-Hara Y, Hara KY, Yamakawa-Kobayashi K. Evolution of a horizontally acquired fatty acid desaturase enables the biosynthesis of omega-3 polyunsaturated fatty acids in Collembola.
Insect Biochemistry and Molecular Biology, 183: 104381 (2025). - Ohhara, Y*., Sagisaka, C., Yamakawa-Kobayashi, K. The collembolan Sinella dubiosa produces eicosapentaenoic acid.
Comp Biochem Physiol B Biochem Mol Biol. 269:110900 (2023). - Twining C*. W., Bernhardt J. R., Derry A. M., Hudson C. M., Ishikawa A., Kabeya N., Kainz M. J., Kitano J., Kowarik C., Ladd S. N., Leal M. C., Scharnweber K., Shipley J. R., Matthews B. The evolutionary ecology of fatty-acid variation: Implications for consumer adaptation and diversification. Ecology letters, 24(8), 1709–1731 (2021).
この記事を書いた人

-
静岡県立大学 食品栄養科学部 食品生命科学科 助教。
2014年 静岡県立大学大学院 生活健康科学研究科 博士後期課程修了。岡崎統合バイオサイエンスセンターおよびカリフォルニア大学リバーサイド校での博士研究員を経て、2015年10月より現職。おもに「虫」と「油」の研究を行っており、虫たちの発育を支える脂質シグナルの分子実体や、脂肪酸代謝の多様性やその進化的意義を垣間見たいと思っています。また、昆虫やトビムシを水産飼料として利用するための基礎研究にも従事しています。
この投稿者の最近の記事
 研究成果2025年10月28日「陸のプランクトン」トビムシに秘められたオメガ3脂肪酸合成経路の発見
研究成果2025年10月28日「陸のプランクトン」トビムシに秘められたオメガ3脂肪酸合成経路の発見


