外環境を感知してエネルギー源を切り替えるエコロジカルなナノモーター
くるくる回る細菌の運動器官 – べん毛
私たちの身の回りには、肉眼では見えない生物、「微生物」がありとあらゆる場所にいます。一般に微生物というと、近年多発する食中毒事件によって、「病原菌」のような悪いイメージを持っている方が多いかもしれません。しかし、醤油や味噌を始めとする発酵食品や多くの抗生物質などは微生物が作り出すものであり、私たちはその恩恵を受けています。また、生態系の最終分解者として彼らとは切っても切れない関係です。
微生物の大きなカテゴリーのなかに、細菌と呼ばれる一群があります。一般的に知られている細菌は、大腸菌やサルモネラ菌、納豆菌でしょうか? 彼らの多くは運動器官として、「べん毛」と呼ばれる細い長いらせん状の繊維を菌体表面から生やしています。そして、あたかも船のスクリュープロペラのように回転させることによって推進力を得て、さまざまな環境を移動することができます。
べん毛の根元には、人工の回転モーターと非常によく似た形の回転モーターが存在します。べん毛モーターは、大まかに回転子と固定子と呼ばれる部分から構成され、これらの相互作用によってモーターが回転します。モーターのエネルギー源は細胞の外から内に流れる水素イオン(H+)もしくはナトリウムイオン(Na+)で、固定子はこのエネルギーを回転力に変換します。その変換効率はほぼ100%で、しかもF1レースカーのエンジンよりも高速回転できます。もちろん、これらすべてタンパク質でできており、べん毛モーターはまさにナノスケールの超精密で高性能な生体分子機械なのです。
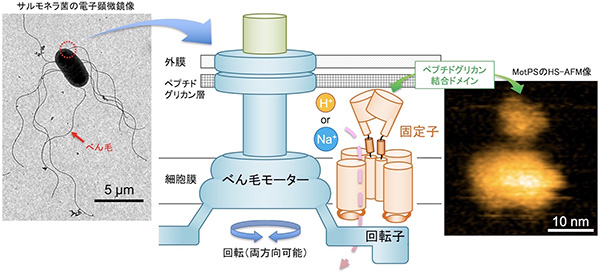
イオンを流すだけじゃない! – べん毛モーター固定子
固定子はイオンを流すことから、イオンチャネルと呼ばれるグループに属します。イオンチャネルは、イオンが流れる通り道とその出入り口を開閉制御する部分から構成されます。しかし固定子の場合、これら以外に細胞膜の外側を囲む細胞壁のペプチドグリカン層に結合する部分、ペプチドグリカン結合(PGB)ドメインを持っています。以前から、固定子はバイオセンサーとして働き、このPGBドメインが外環境の変化を感知してモーターの周りに配置される固定子の数を制御すると考えられていました。しかし、そのしくみはまったく謎でした。
枯草菌べん毛モーターの固定子であるMotPS複合体はNa+で動作します。MotPS複合体はNa+が存在するとべん毛モーターの周りに集まることができます。その仕組みを理解するため、私たちはMotPS複合体の単離精製に挑戦しました。固定子は膜タンパク質複合体であることからその精製は難しく、さらに複合体の状態のまま精製するのは大変困難な課題でしたが、多くの条件検討の結果、複合体構造を保ったまま単離精製することに成功しました。
次に、MotPS複合体がどのようにしてNa+を感知するのかを知るため、高時間高空間分解能で生体分子の観察が可能な高速原子間力顕微鏡(HS-AFM)を活用しました。HS-AFMは、カンチレバーと呼ばれる鋭い探針で試料表面をなぞることで試料表面の凹凸を高分解能で可視化できる顕微鏡で、水溶液中での生体分子の反応や構造変化をリアルタイムで観察できます。
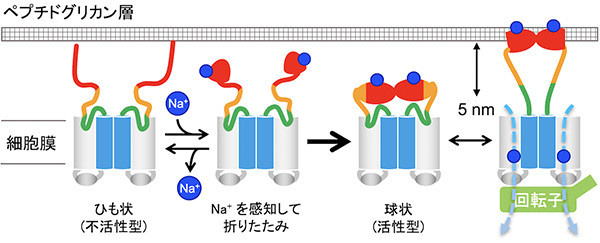
単離精製したMotPS複合体をHS-AFMで観察したところ、大きな楕円状の細胞膜貫通領域と小さな楕円状のPGBドメインが細い糸(アミノ酸の鎖)で繋がった分子形態が明らかとなりました。さらに、PGBドメインは観察時の溶液中にNa+が存在するときのみ球状構造を形成し、Na+が存在しない時は立体構造が解きほぐされてひも状になることを発見しました。そして再びNa+を添加すると折りたたまれて球状構造に戻るという、可逆的な反応であることもわかりました。
これらの結果から、MotPS複合体ではNa+の結合解離によって、PGBドメインが機能状態から機能しない状態へと可逆的に変化することが明らかになりました。MotPS複合体はこのような仕組みによって、Na+が環境中に存在する時にのみモーターに組み込まれることができるのです。

掴めるか!? – モーターへの組み込み機構
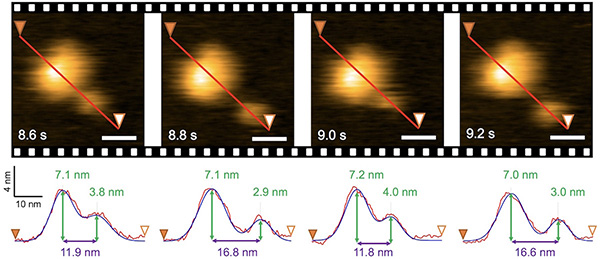
固定子複合体は膜タンパク質なので、数多くが細胞膜中をプカプカ漂っています。そして、モーターに組み込まれて機能するときのみ、球状構造を形成したPGBドメインが細胞壁であるペプチドグリカン層に結合すると考えられます。しかし、細胞膜とペプチドグリカン層のあいだにはおよそ5 nmの間隙があります。私たちはHS-AFM観察によって、MotPSの膜貫通領域(大きな球状構造)とPGBドメイン(小さな球状構造)との距離がダイナミックに変化することを見出しました。
そこで、大小2つの球状構造の重心間距離を精密に測定したところ、およそ12-17 nmの範囲でダイナミックに変化していました。つまり、大小2つの球状構造を繋ぐ細い糸が最大で5 nmの範囲で伸びたり縮んだりするのです。これは、PGBドメインがペプチドグリカン層に結合するために必要な距離5 nmと一致します。これらの結果は、MotPS複合体がモーターに結合すると、PGBドメインが5 nm伸びてペプチドグリカン層を掴み、その結果MotPS複合体がNa+駆動型の固定子として働くができることを示唆しています。
おわりに – 究極の環境性能へ
細菌べん毛モーターはおよそ30種類のタンパク質から作られる超分子複合体のナノマシンで、エネルギー変換効率はほぼ100%と概算され、大変高性能なモーターとして機械工学や医療の分野で大変注目を集めています。その心臓部である固定子は、モーターを回転させるために必要なエネルギー源があるときのみ活性化してモーターに組み込まれるというように、「今、無駄なく使えるエネルギー」を選択していることが明らかになりました。私たち人類の科学力ではまだ人工的にナノマシンを作ることはできませんが、細菌べん毛モーターからこのようなエコロジカルな観点も視野に入れたナノテク技術を学ぶことができます。本研究はべん毛モーターの回転機構の解明への大きな第一歩となるだけなく、イオン流エネルギーを利用する高効率で環境に優しい人工モーターへの応用が期待されます。
参考文献
この記事を書いた人

-
寺原直矢(写真左)
日本学術振興会特別研究員(DC2)を経て、2008年東洋大学大学院生命科学研究科修了:博士(生命科学)。2009年東洋大学バイオナノエレクトロニクス研究センター研究助手、2010年大阪大学大学院生命機能研究科にて日本学術振興会特別研究員(PD)。2013年同特任研究員を経て、2016年同特任助教。現在、クライオ電子顕微鏡による固定子複合体の高分解能構造解析を進める。
難波啓一(写真中央)
1980年大阪大学大学院基礎工学研究科物理系専攻博士課程修了(工学博士)。学術振興会奨励研究員、米国Brandeis大学、Vanderbilt大学博士研究員を経て、86年より科学技術振興事業団ERATO超分子柔構造プロジェクトグループリーダー、92年より松下電器国際研究所リサーチディレクター、2002年より大阪大学大学院生命機能研究科教授、そして2017年より特任教授。1997〜2003年は科学技術振興事業団ERATOプロトニックナノマシンプロジェクト総括責任者、2003〜08年は科学技術振興機構ICORP超分子ナノマシンプロジェクト研究総括を兼任。専門分野は生物物理学、構造生物学で、べん毛モーターやアクトミオシンモーターの構造と動態から力発生・エネルギー変換メカニズムの解明をめざしている。
南野徹(写真右)
1997年広島大学大学院生物圏科学研究科修了、1997〜2000年エール大学博士研究員、2000〜2002年難波ERATOプロジェクト研究員、2003〜2005年難波ICORPプロジェクトグループリーダー、2005年大阪大学大学院生命機能研究科助手を経て、2010年より現職。べん毛蛋白質輸送装置の選択的物質輸送メカニズムやエネルギー変換メカニズムの研究に従事するとともに、べん毛モーターの力学応答とそれに伴うモーターの再編成の研究にも従事。



