細胞内タンパク質分解を担う「オートファゴソーム」のサイズ制御 – カギとなるのは小胞体膜タンパク質ERdj8
オートファゴソームのサイズはどう調節されているのか?
オートファジーとは、細胞の内部で、タンパク質をはじめとする自身の構成成分を分解する仕組みです。細胞質成分を、二重の膜構造であるオートファゴソームと呼ばれる球状のオルガネラで包み込みます。オートファゴソームはタンパク質分解酵素などさまざまな加水分解酵素を多く含むリソソームと融合することで、分解基質が分解されます。
オートファジーにおいては無作為に基質を包み込むだけではなく、ダメージを受けたミトコンドリアを分解するマイトファジーや細胞に侵入してきた細菌を分解するゼノファジーなどの選択的オートファジーも知られています。1990年代、筆者(野田)らがかつて所属した大隅良典博士の研究室や海外の研究グループにおいて、オートファゴソーム形成を担うAtg遺伝子群が同定されてから、オートファゴソームを形成する仕組みは徐々に理解されてきました。
興味深いことに、非選択的なオートファジーにおけるオートファゴソームの直径は1μm程度でほぼ一定であるのに対して、ミトコンドリアや細菌などの大きな基質を包み込む際には3μmを超える大きなオートファゴソームが形成されることもあります。しかしながら、オートファゴソームのサイズがどのように調節されているのか、その仕組みについてはほぼ理解されていません。
小胞体とオートファジーの密接な連携の仕組み
小胞体は細胞内における細胞内小器官オルガネラの一種であり、細胞内を張り巡らせるようなネットワーク構造をとっています。分泌タンパク質や膜タンパク質の合成、脂質の合成、カルシウムの貯蔵など、細胞を維持するうえで重要な働きをしています。
近年、顕微鏡技術が進歩し小胞体の詳細な構造が徐々に明らかにされつつあります。小胞体は均一な構造ではなく、特定の領域(小胞体サブドメイン)が存在することが明らかになってきました。
オートファゴソームの形成は小胞体の近傍で起こることが知られてますが、最近では小胞体膜に存在するリン脂質合成酵素が、局在化した小胞体サブドメインの近傍において起こることが報告されています。リン脂質を成分としているオートファゴソーム膜の形成で脂質合成が起こることは理解しやすいと思います。
ERdj8によるオートファゴソームのサイズ制御は線虫からヒトまで共通
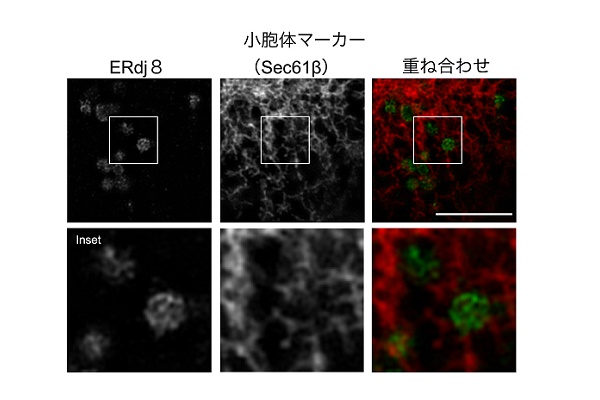
今回、我々が注目したERdj8は、小胞体に存在する膜タンパク質です。ERdjファミリータンパク質はこれまでに7種類報告されており、分泌タンパク質や膜タンパク質の折りたたみや構造異常タンパク質の分解、Ca2+の濃度の調節に関わることが知られています。
ERdj8はERdjファミリータンパク質の8番目にあたりますが、その機能はまったくわかっていませんでした。ヒト由来のHeLa細胞において超解像顕微鏡を用いて観察したところ、ERdj8は小胞体ネットワーク上のごく一部に局在していました。その近傍にはオートファゴソーム形成に関わるAtgタンパク質やリン脂質合成酵素であるPISが存在していました。
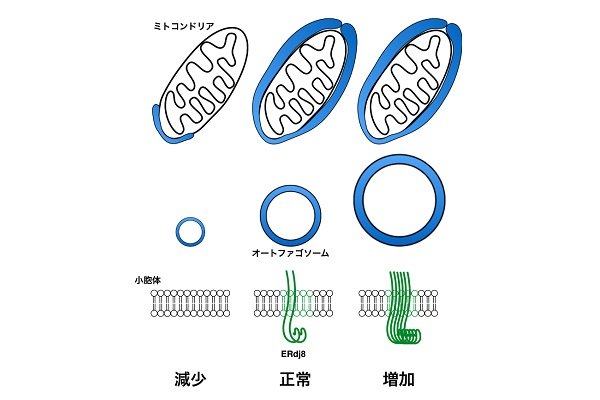
ERdj8を過剰に発現させると、オートファゴソームのサイズが大きくなりました。ERdj8の発現を抑制すると、オートファゴソームのサイズが小さくなり、ミトコンドリアや大きなラテックスビーズなどを包めなくなりましたが、小さなオートファジーの基質は包むことができました。
線虫のERdj8ホモログであるdnj-8の発現を抑制すると、筋細胞中のミトコンドリアの分解は阻害されたのに対し、小型のミトコンドリアである父性ミトコンドリアの分解には影響を及ぼしませんでした。これらの結果は生物種間を超えて、ERdj8がオートファゴソームの大きさを制御するうえで重要な役割を担っていることを示しています。
ERdj8の機能解析からオートファゴソームのサイズ制御機構の解明へ
今回発表した論文では、オートファゴソームの大きさを変化させる際にはERdj8が重要な役割を担うという現象を明らかにしました。ERdj8が小胞体のリン脂質合成酵素が集積するサブドメインと近接して存在することや、ERdj8の機能ドメインがオートファゴソームの大きさの変化に影響を与えること、などをヒントにして、ERdj8がどのようにオートファゴソーム膜の大きさを制御変化させているのか、その分子メカニズムの解析を行っていきたいと思います。
参考文献
Yo-hei Yamamoto#, Ayano Kasai#, Hiroko Omori, Tomoe Takino, Munechika Sugihara, Tetsuo Umemoto, Maho Hamasaki, Tomohisa Hatta, Tohru Natsume, Richard I. Morimoto, Ritsuko Arai, Satoshi Waguri, Miyuki Sato, Ken Sato, Shoshana Bar-Nun, Tamotsu Yoshimori, Takeshi Noda* and Kazuhiro Nagata* “ERdj8 governs the size of autophagosomes during the formation process” J Cell Biol. 2020 Aug 3; 219(8): e201903127. doi: 10.1083/jcb.201903127.
*共同責任著者 #同程度貢献
この記事を書いた人

-
山本 洋平(画像左)
大阪大学 大学院歯学研究科 口腔科学フロンティアセンター 助教。
奈良先端科学技術大学院大学バイオサイエンス研究科動物細胞工学研究室の河野憲二教授のもとで学位取得。その後、京都産業大学タンパク質動態研究所の永田和宏研究室にて博士研究員として研究を行う。2018年1月より現職。学生時代から一貫して、小胞体局在型DnaJタンパク質の研究を続けている。現在はDnaJタンパク質であるERdj8を軸にオートファゴソームのサイズ調節機構に興味を持ち研究を行っている。これからも一途に小胞体分子シャペロンが制御する生命現象を明らかにすることを目標に研究を続けていきたい。
野田 健司(画像右)
大阪大学 大学院歯学研究科 口腔科学フロンティアセンター 教授。
東京大学教養学部基礎科学科第一卒、東京大学大学院理学系研究科相関理化学専攻において大隅良典博士が立ち上げたばかりの研究室において、オートファジー研究の立ち上げに関わる。博士号取得後、大隅研が愛知県岡崎市の基礎生物学研究所への移動に伴い、助手として、学生時代を加えて計13年間その研究を支えてきた。米国留学をはさみ、大阪大学微生物病研究所へ吉森保博士が立ち上げた研究室の助教授として着任後、生命機能研究科へ異動。2012年大阪大学大学院歯学研究科において、教授として独立。オートファジーの分子機構に加えて、新たな細胞生物学の分野の開拓を目指している。