マイコプラズマ・モービレの滑走運動メカニズム – 「あし」がつかんで、ひっぱり、はなす
モリクテス綱とその運動能
モリクテス(Mollicutes)綱は、マイコプラズマ(Mycoplasma)属やスピロプラズマ(Spiroplasma)属などから構成される小さな細菌の小さな集団です。培養可能な生物のなかでもっとも小さなゲノムを持つことや、細菌のなかで唯一、細胞壁であるペプチドグリカン層を持たないことなど、他の細菌とは一線を画する特徴を持っています。
かつてはこれらの特徴が、原始の細胞に近いことを示しているとされていました。しかし本当は逆で、納豆菌やボツリヌス菌に近縁の細菌が、動植物に寄生することで、進化を遂げて生まれたのがモリクテス綱細菌であることが今ではわかっています。
モリクテス綱細菌には寄生して生きていくためのいくつかの能力が備わっていますが、そのなかの際立ったものが、グループ全体で3種類もの運動能を持つことです。地球上の全生命でこれまでに見つかった細胞のすべての運動能が18種類であることを考えると、3種類は驚きの数字です。
一般に細菌の運動はペプチドグリカン層を必要としますから、モリクテス綱細菌がペプチドグリカン層を捨てて動けなくなった後に、宿主の免疫細胞から逃げのびなければいけないという、進化上の強い選択圧がかかって新たな運動能を獲得したものと考えられます。
マイコプラズマ・モービレの滑走運動
コイ科の観賞魚、テンチ(Tench)のえらの病変から単離されたマイコプラズマ・モービレ(Mycoplasma mobile)はその名のとおり、高速で動き回る種類です。私たちは略して「モービレ」と呼んでいます。
マイコプラズマ・モービレ滑走運動のリアルタイムビデオ。細胞長は平均で0.8 μm。
モービレは菌体の片側に滑走装置と呼ばれる突起を形成し、その装置で動物細胞やガラスなどの固形物表面にはりつき、すべるように動く滑走運動を行います。滑走の速さは、毎秒4.5 μm(マイクロメートル)にも達しますが、菌体の長さが約0.8 μmですから、人間の比率に直すと毎秒9.6 mの速さで走っていることになります。ビデオを見るとスイスイと動くさまがコミカルでも、楽しそうでもあります。
モービレの滑走装置とは?
私たちは1997年にこのモービレ滑走運動のメカニズム解明の研究を始めました。なぜなら、この運動能がその当時によく研究されていた筋肉や、真核生物の鞭毛や、細菌のべん毛などとはまったく異なるものと考えられたからです。その予想が正しかったことは、私たちの研究で明らかになりました。
滑走装置は、細胞膜外側表面の構造と内側の構造に分けることができます。表面には3種類の巨大タンパク質が同じ数ずつ整然と並んでいます。3種類のタンパク質の役割はそれぞれ、宿主細胞表面の構造をつかむ「あし」、内部構造中のモーターが発生した動きを伝達・拡大する「クランク」、これらのタンパク質を固定する「ボルト」、の役割を果たしています。特に、あしとクランクのタンパク質は、役割もですが分子そのものがそれらしい形状をしています。寄生性の細菌の多くが宿主細胞にしがみつくための接着タンパク質を持っているのですが、あしのタンパク質はモービレの祖先が持っていた接着タンパク質から進化したものと考えられます。
内部構造はクラゲのような形状です。驚いたことにクラゲの触手を構成しているタンパク質の一部は、ATP合成酵素の触媒サブユニットと30%~40%ものアミノ酸配列の相同性、すなわち構造の類似性があります。ATP合成酵素は細菌の細胞膜やミトコンドリアの内膜に存在し、細胞の呼吸により形成されたプロトン(H+)駆動力によりATPを合成する装置です。膜を横切ろうとするプロトンが中を通るときに装置は回転し、回転に伴う構造変化をもとにATPを合成します。また、逆にATPを消費することでプロトンを輸送することもできます。
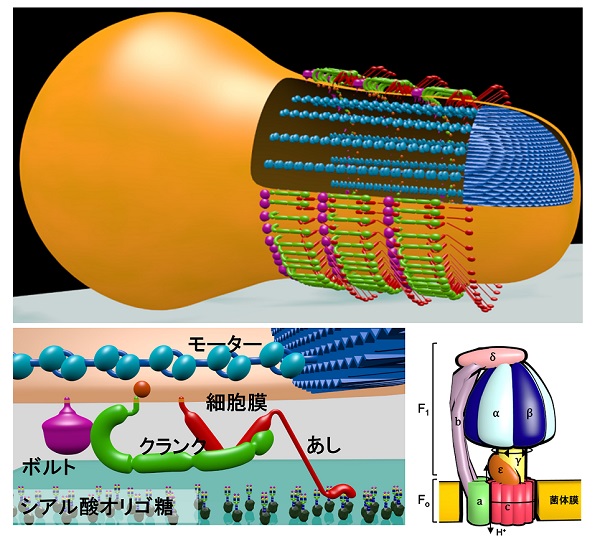
(c) Miyata et al., Curr. Opin. Microbiol. 29, 15-21 (2016)
滑走装置内部構造とこのATP合成酵素の構造が似ているということは、滑走装置の起源が回転するATP合成酵素と接着タンパク質の出会いから形成されたことを意味しています。実際に、細胞膜に界面活性剤(洗剤)で穴をあけて細胞の中身を流し出してしまうと、細胞は固形物表面にはりついたまま止まってしまいますが、そこにATPを加えるともとと同じように滑走しました。私たちはこの現象を「滑走ゴースト」と命名しました。この発見から、ATPの加水分解が滑走運動に直接関わっていることがわかりました。次に、この内部構造で発生した動きがどのように「あし」に伝わるかが謎になりました。
滑走ゴースト実験のリアルタイムビデオ。界面活性剤を入れると止まり、ATPを入れると再び滑走する。
内部構造から「あし」へ動きの伝達
私たちは、滑走ゴーストを凍結し、その電子顕微鏡像を数百の異なった角度から撮影しました。それらの像からCTスキャンと同じ方法で元の構造を再構築しました。その結果、滑走装置の内部構造が膜を裏打ちするように配置されており、菌体外部の構造へとつながっていることが明らかになりました。
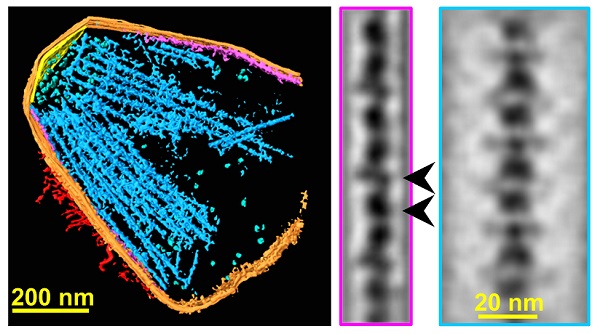
(c) Nishikawa et al., mBio. 10(6). e02846-19. (2019)
また、ATP加水分解各段階における滑走装置の内部構造と、滑走のあしのその結合対象であるシアル酸オリゴ糖への結合を調べました。シアル酸オリゴ糖は動物細胞表面に多く存在する構造で、インフルエンザウイルスの結合対象としても知られています。これらの実験の結果、ATPの加水分解に伴いモーターの形が変化していること、それに伴って「あし」がシアル酸オリゴ糖をつかんだりはなしたりしていることが明らかになりました。
そうして、私たちは滑走運動メカニズムの新しいモデルを提案しました(下図)。驚いたことに、そのメカニズムにおけるATP加水分解と足場のひっぱりとの組み合わせは、まったく異なった構造を持つヒトを含む動物の筋肉の収縮メカニズムとよく類似したものでした。
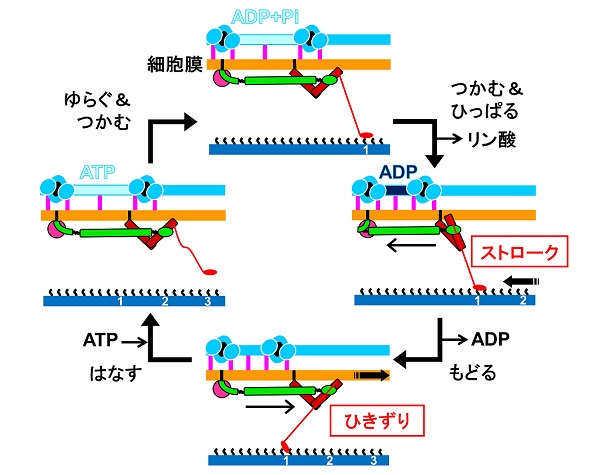
(c) Nishikawa et al., mBio. 10(6). e02846-19. (2019)
私たちはこれから、この、世にも珍しい「運動超分子マシナリー」ともいえる滑走装置がどのような構造を持ち動くのか、そしてどのように進化してきたかを原子レベルで解明し、生命の本質であるエネルギー変換と生き残りの秘密に迫っていきます。
参考文献
1. 地球上のすべての生命の細胞運動能をはじめて俯瞰した論文
Miyata M*, Robinson RC, Uyeda T Q P, Fukumori Y, Fukushima S, Haruta S, Homma M, Inaba K, Ito M, Kaito C, Kato K, Kenri T, Kinosita Y, Kojima, Minamino T, Mori H, Nakamura S, Nakane D, Nakayama K, Nishiyama M, Shibata S, Shimabukuro K, Tamakoshi M, Taoka A, Tashiro Y, Tulum I, Wada H, and Wakabayashi K. (2020) “Tree of motility – A proposed history of motility systems in the tree of life.” Genes to Cells. 25(1):6-21. doi: 10.1111/gtc.12737.
2. モービレ運動能の研究をまとめた論文
Miyata M* and Hamaguchi T (2016) “Prospects for the gliding mechanism of Mycoplasma mobile.” Current Opinion in Microbiology. 29, 15-21. doi: 10.1016/j.mib.2015.08.010.
3. 内部構造からあしへ動きの伝達を明らかにした論文
Nishikawa MS, Nakane D, Toyonaga T, Kawamoto A, Kato T, Namba K, and Miyata M* (2019) “Refined mechanism of Mycoplasma mobile gliding based on structure, ATPase activity, and sialic acid binding of machinery.” mBio. 10(6). e02846-19. doi: 10.1128/mBio.02846-19.
*責任著者
この記事を書いた人

-
宮田 真人(みやた まこと)
大阪市立大学大学院理学研究科・生物地球系専攻・教授。
1983年大阪大学理学部生物学科卒業。1988年同大学院修了、理学博士。1988年大阪市大理学部助手。講師、助教授を経て2006年より現職。1997年ウイーン獣医大学・訪問科学者。2000~2001年ローランド科学研究所およびハーバード大学・訪問学者。2003~2007年科学技術振興機構(JST)さきがけ研究者。2012~2017年文科省科研費・新学術領域「運動超分子マシナリーが織りなす調和と多様性」領域代表。2019年よりJST CREST研究「合成細菌JCVI syn3.0Bとゲノム操作を用いた細胞進化モデルの構築」研究代表者。